
Какое свойство обеспечивает процесс редупликации днк

Молекула ДНК — это находящаяся в хромосоме структура. Одна хромосома содержит одну такую молекулу, состоящую из двух нитей. Редупликация ДНК — это передача информации после самовоспроизведения нитей от одной молекулы на другую. Она присуще как ДНК, так и РНК. В данной статье рассматривается процесс редупликации ДНК.

Общие сведения и виды синтеза ДНК
Известно, что нити в молекуле закручены. Однако, когда начинается процесс редупликации ДНК, они деспирализуются, затем отходят в стороны, и на каждой синтезируется новая копия. По завершении появляются две абсолютно идентичные молекулы, в каждой из которых присутствует материнская и дочерняя нити. Такой синтез получил название полуконсервативный. Молекулы ДНК отодвигаются, оставаясь при этом в единой центромере, и окончательно расходятся лишь тогда, когда у этой центромеры начинается процесс деления.

Другой вид синтеза получил название репаративный. Он, в отличие от предыдущего, не связан с какой-либо клеточной стадией, но начинается при возникновении повреждений ДНК. Если они носят слишком обширный характер, то клетка в конце концов погибает. Однако, если повреждения локальны, то их можно восстановить. В зависимости от проблемы восстановлению подлежит отдельная или две сразу цепочки ДНК. Этот, как его еще называют, внеплановый синтез не занимает продолжительного времени и не требует больших энергозатрат.
Но когда происходит редупликация ДНК, то расходуется много энергии, материала, продолжительность его растягивается на часы.
Редупликация делится на три периода:
- инициацию;
- элонгацию;
- терминацию.
Рассмотрим подробнее эту последовательность редупликации ДНК.
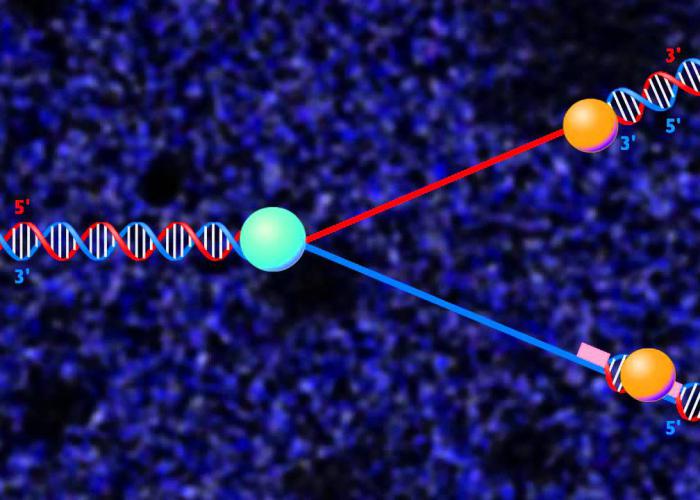
Инициация
В ДНК человека — несколько десятков миллионов пар нуклеотидов (у животных их насчитывается всего сто девять). Редупликация ДНК начинается во многих местах цепочки по следующим причинам. Примерно в это же время в РНК происходит транскрипция, но на время синтеза ДНК она приостанавливается в некоторых отдельных местах. Поэтому перед таким процессом в цитоплазме клетки накапливается достаточное количество вещества для того, чтобы поддержать экспрессию генов и чтобы жизнедеятельность клетки не была нарушена. Ввиду этого процесс должен проходить как можно быстрее. Трансляция в этот период осуществляется, а транскрипция не ведется. Как показали исследования, редупликация ДНК происходит сразу в нескольких тысячах точек — небольших участках с определенной последовательностью нуклеотидов. К ним присоединяются специальные инициаторные белки, к которым в свою очередь присоединяются другие ферменты редупликации ДНК.
Фрагмент ДНК, где происходит синтез, называется репликоном. Он начинается от точки начала и заканчивается тогда, когда фермент завершает репликацию. Репликон автономен, а также снабжает весь процесс собственным обеспечением.
Процесс может начаться не со всех точек сразу, где-то он начинается раньше, где-то — позже; может протекать в одном или в двух противоположных направлениях. События происходят в следующем порядке, когда образуются:
- репликационная вилка;
- РНК-затравка.
Репликативная вилка
Эта часть представляет собой процесс, при котором на отсоединенных нитях ДНК происходит синтез дезоксирибонуклеиновых нитей. Вилки при этом образуют так называемый глазок редупликации. Процессу предшествует целый ряд действий:
- освобождение от связи с гистонами в нуклеосоме — такие ферменты редупликации ДНК как метилирование, ацетилирование и фосфорилирование производят химические реакции, в результате которых белки теряют свой положительный заряд, что способствует их высвобождению;
- деспирализация — это раскручивание, которое необходимо для дальнейшего освобождения нитей;
- разрыв связей водорода между нитями ДНК;
- их расхождение в разные стороны молекулы;
- фиксация, происходящая при помощи белков SSB.
РНК-затравка
Синтез осуществляет фермент, под названием ДНК-полимераза. Однако начать его самостоятельно он не может, поэтому это делают другие ферменты — РНК-полимеразы, которые называют еще РНК-затравками. Они синтезируются параллельно дезоксирибонуклеиновым нитям по комплементарному принципу. Таким образом, инициация заканчивается синтезом двух РНК-затравок на двух разорванных и отошедших в разные стороны нитях ДНК.
Элонгация

Данный период начинается с присоединения нуклеотида и 3′ концу РНК-затравки, что осуществляет уже упомянутая ДНК-полимераза. К первому она присоединяет второй, третий нуклеотид, и так далее. Основания новой нити соединяются с материнской цепочкой водородными связями. Считается, что синтез нити идет в направлении 5 ‘- 3’.
Там, где он происходит в сторону репликационной вилки, синтез протекает непрерывно и удлиняется при этом. Поэтому такую нить называют ведущей или лидирующей. На ней РНК-затравки больше не формируются.
Однако на противоположной материнской нити ДНК-нуклеотиды продолжают присоединяться к РНК-затравке, и дезоксирибонуклеиновая цепь синтезируется в противоположном от вилки редупликации направлении. Ее в этом случае называют запаздывающей или отстающей.
На отстающей нити синтез происходит фрагментарно, где по окончании одного участка начинается синтез на другом участке поблизости при помощи все той же РНК-затравки. Таким образом, на запаздывающей цепи имеются два фрагмента, которые соединены ДНК и РНК. Они получили название фрагменты Оказаки.
Далее все повторяется. Тогда расплетается другой виток спирали, разрываются связи водорода, нити расходятся в стороны, ведущая цепь удлиняется, на отстающей синтезируется следующий фрагмент РНК-затравки, после чего — фрагмент Оказаки. После этого на запаздывающей нити РНК-затравки разрушаются, а фрагменты ДНК соединяются в одну. Так на этой цепи происходит одновременно:
- образование новых РНК-затравок;
- синтез фрагментов Оказаки;
- разрушение РНК-затравок;
- воссоединение в одну единую цепь.
Терминация
Процесс продолжается до тех пор, пока две репликативные вилки не встретятся, или одна из них не подойдет к концу молекулы. После встречи вилок дочерние нити ДНК соединяются ферментом. В случае же, если вилка отошла к концу молекулы, редупликация ДНК заканчивается с помощью специальных ферментов.
Коррекция
В данном процессе важная роль отводится контролю (или коррекции) редупликации. К месту синтеза поступают все четыре вида нуклеотидов, а путем пробного спаривания ДНК-полимераза отбирает те, которые необходимы.
Нужный нуклеотид должен быть способен сформировать столько же связей водорода, сколько аналогичный нуклеотид на матричной нити ДНК. Кроме того, между сахарофосфатными остовами должно быть определенное постоянное расстояние, соответствующее трем кольцам в двух основаниях. Если нуклеотид не соответствует этим требованиям, соединение происходить не будет.
Контроль проводится перед включением его в состав цепи и перед включением последующего нуклеотида. После этого формируется связь в остове сахарофосфата.
Мутационная изменчивость
Механизм редупликации ДНК, несмотря на высокий процент точности, всегда имеет нарушения в нитях, называющихся в основном «генными мутациями». Примерно на тысячу нуклеотидных пар приходится одна ошибка, которая называется конвариантная редупликация.
Она случается по разным причинам. К примеру, при высокой или слишком низкой концентрации нуклеотидов, дезаминирования цитозина, присутствия мутагенов в области синтеза, и другое. В некоторых случаях ошибки могут исправиться репарационными процессами, в других исправление становится невозможным.
Если повреждение коснулось неактивного места, ошибка не будет иметь тяжелых последствий, когда происходит процесс редупликации ДНК. Последовательность нуклеотида того или иного гена может проявиться с ошибкой спаривания. Тогда дело обстоит иначе, и негативным результатом может стать как гибель этой клетки, так и гибель всего организма. Следует также учитывать, что генные мутации основаны на мутационной изменчивости, которая делает генофонд пластичнее.
Метилирование

В момент синтеза или сразу после него происходит метилирование цепей. Считается, что у человека этот процесс нужен для того, чтобы сформировать хромосомы и регулировать транскрипцию генов. В бактериях данный процесс служит защитой ДНК от разрезания его ферментами.
Источник
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 7 марта 2020; проверки требуют 4 правки.
У этого термина существуют и другие значения, см. Репликация.
Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК-полимераза (Polα), (4) ДНК-лигаза, (5) РНК-праймер, (6) праймаза, (7) фрагмент Оказаки, (8) ДНК-полимераза (Polδ), (9) хеликаза, (10) белки, связывающие одноцепочечную ДНК, (11) топоизомераза
Процесс репликации: раскручивание двойной спирали ДНК — синтез комплементарных цепей ДНК-полимеразой — образование двух молекул ДНК из одной
Реплика́ция (от лат. replicatio — возобновление) — процесс создания двух дочерних молекул ДНК на основе родительской молекулы ДНК. Репликацию ДНК осуществляет сложный комплекс, состоящий из 15—20 различных белков-ферментов, называемый реплисомой (англ.)[1] С помощью специальных ферментов двойная спираль материнской ДНК расплетается на две нити, на каждой образовавшейся нити достраивается вторая нить, образуя две идентичных дочерних молекулы ДНК, которые затем скручиваются в отдельные спирали. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение.
История изучения[править | править код]
Каждая молекула ДНК состоит из одной цепи исходной родительской молекулы и одной вновь синтезированной цепи. Такой механизм репликации называется полуконсервативным. В настоящее время этот механизм считается доказанным благодаря опытам Мэтью Мезельсона и Франклина Сталя (1958 г.)[2]. Ранее существовали и две другие модели: «консервативная» — в результате репликации образуется одна молекула ДНК, состоящая только из родительских цепей, и одна, состоящая только из дочерних цепей; «дисперсионная» — все получившиеся в результате репликации молекулы ДНК состоят из цепей, одни участки которых вновь синтезированы, а другие взяты из родительской молекулы ДНК. Молекула ДНК разрезается пополам и образутся два шаблона. Два шаблона выходят из репликационной вилки. Если представить их в выпрямленном виде, то можно видеть линейку из гребёнок, которые соединены концами, но имеют промежуток. Представим что одна гребёнка синяя, а другая — красная. Теперь подставим нижнюю красную (она из пяти гребней, как и верхняя) пятым концом к третьему верхнему (третьей верхней иголке). Удлиним цепь и сверху и снизу. Как бы получится: пять, три, пять и т. д.- наверху и снизу тоже. Потом к этим гребёнкам добавляются после выхода шаблонов (гребёнок) из репликационной вилки ещё два шаблона. Из одной молекулы ДНК получается две идентичные материнской (если нет мутаций) молекулы, это называется полуконсервативностью.
Общие представления[править | править код]
Репликация ДНК — ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа:
- инициация репликации
- элонгация
- терминация репликации.
Регуляция репликации осуществляется в основном на этапе инициации. Это достаточно легко осуществимо, потому что репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. В геноме таких сайтов может быть как всего один, так и много. С понятием сайта инициации репликации тесно связано понятие репликон. Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномы бактерий, как правило, представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации. Геномы эукариот (а также их отдельные хромосомы) состоят из большого числа самостоятельных репликонов, это значительно сокращает суммарное время репликации отдельной хромосомы. Молекулярные механизмы, которые контролируют количество актов инициации репликации в каждом сайте за один цикл деления клетки, называются контролем копийности. В бактериальных клетках помимо хромосомной ДНК часто содержатся плазмиды, которые представляют собой отдельные репликоны. У плазмид существуют свои механизмы контроля копийности: они могут обеспечивать синтез как всего одной копии плазмиды за клеточный цикл, так и тысяч копий[1].
Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более распространена двунаправленная репликация. Через некоторое время после начала репликации в электронный микроскоп можно наблюдать репликационный глазок — участок хромосомы, где ДНК уже реплицирована, окружённый более протяжёнными участками нереплицированной ДНК[1].
В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500—5000 — у эукариот[3].
| Фермент | Функция |
|---|---|
| ДНК-гираза | Вносит временные двуцепочечные разрывы в ДНК, облегчая её разматывание. |
| Хеликаза | Разделяет цепи двухцепочечной молекулы ДНК на одинарные цепи. |
| SSB-белки | Связывают одноцепочечные фрагменты ДНК и предотвращают комплементарное спаривание. |
| Праймаза | Синтезирует РНК-затравку (праймер) — короткий фрагмент РНК, которая является инициатором в работе ДНК-полимеразы (полимераза не способна синтезировать ДНК с нуля, но может добавлять нуклеотиды к уже имеющимся). |
| ДНК-полимераза | Синтезирует ДНК, связываясь с праймером. Следует отметить, что один конец материнской ДНК полимераза синтезировала непрерывно и в одном направлении, а второй — в противоположном — фрагментами. |
| Белки скользящего зажима (застежки) | Окружают кольцом ДНК и «скользят» по ней вместе с продвигающейся вперед фермента ДНК-полимеразы. Они предотвращают диссоциацию фермента от матрицы ДНК и повышают эффективность его работы. |
| РНКаза H | Удаляет уже ненужные фрагменты РНК-затравки. |
| ДНК-лигаза | Сшивает фрагменты ДНК (фрагменты Оказаки). |
| Теломераза | Добавляет особые повторяющиеся последовательности нуклеотидов к одному концу цепи ДНК на участках теломер, тем самым компенсируя их укорачивание во время деления. |
| Реплисома (комплекс всех ферментов репликации) | Движется вдоль молекулы ДНК-матрицы, расплетая её и наращивая комплементарные цепи ДНК. |
Молекулярный механизм репликации[править | править код]
Ферменты (хеликаза, топоизомераза) и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у прокариот[уточнить]
осуществляется несколькими разными ДНК-полимеразами. ДНК-полимераза I действует на запаздывающей цепи для удаления РНК-праймеров и дорепликации очищенных мест ДНК. ДНК полимераза III — основной фермент репликации ДНК, осуществляющий синтез ведущей цепи ДНК и фрагментов Оказаки при синтезе запаздывающей цепи. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный.
Цепи молекулы ДНК расходятся, образуют репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле.
Характеристики процесса репликации
- матричный — последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской цепи в соответствии с принципом комплементарности;
- полуконсервативный — одна цепь молекулы ДНК, образовавшейся в результате репликации, является вновь синтезированной, а вторая — материнской;
- идёт в направлении от 5’-конца новой молекулы к 3’-концу;
- полунепрерывный — одна из цепей ДНК синтезируется непрерывно, а вторая — в виде набора отдельных коротких фрагментов (фрагментов Оказаки);
- начинается с определённых участков ДНК, которые называются сайтами инициации репликации (англ. origin)[4].
Примечания[править | править код]
Литература[править | править код]
- Сохранение ДНК в ряду поколений: Репликация ДНК (Фаворова О.О., СОЖ, 1996)PDF (151 KB)
- Репликация ДНК (анимация) (англ.)
Источник
Репликация ДНК — процесс самоудвоения, главное свойство молекулы ДНК. Репликация относится к категории реакций матричного синтеза, идет с участием ферментов. Под действием ферментов молекула ДНК раскручивается, и около каждой цепи, выступающей в роли матрицы, по принципам комплементарности и антипараллельности достраивается новая цепь. Таким образом, в каждой дочерней ДНК одна цепь является материнской, а вторая — вновь синтезированной. Такой способ синтеза называется полуконсервативным.
«Строительным материалом» и источником энергии для репликации являются дезоксирибонуклеозидтрифосфаты (АТФ, ТТФ, ГТФ, ЦТФ), содержащие три остатка фосфорной кислоты. При включении дезоксирибонуклеозидтрифосфатов в полинуклеотидную цепь два концевых остатка фосфорной кислоты отщепляются, и освободившаяся энергия используется на образование фосфодиэфирной связи между нуклеотидами.
В репликации участвуют следующие ферменты:
- геликазы («расплетают» ДНК);
- дестабилизирующие белки;
- ДНК-топоизомеразы (разрезают ДНК);
- ДНК-полимеразы (подбирают дезоксирибонуклеозидтрифосфаты и комплементарно присоединяют их к матричной цепи ДНК);
- РНК-праймазы (образуют РНК-затравки, праймеры);
- ДНК-лигазы (сшивают фрагменты ДНК).
С помощью геликаз в определенных участках ДНК расплетается, одноцепочечные участки ДНК связываются дестабилизирующими белками, образуется репликационная вилка. При расхождении 10 пар нуклеотидов (один виток спирали) молекула ДНК должна совершить полный оборот вокруг своей оси. Чтобы предотвратить это вращение ДНК-топоизомераза разрезает одну цепь ДНК, что дает ей возможность вращаться вокруг второй цепи.
ДНК-полимераза может присоединять нуклеотид только к 3′-углероду дезоксирибозы предыдущего нуклеотида, поэтому данный фермент способен передвигаться по матричной ДНК только в одном направлении: от 3′-конца к 5′-концу этой матричной ДНК. Так как в материнской ДНК цепи антипараллельны, то на ее разных цепях сборка дочерних полинуклеотидных цепей происходит по-разному и в противоположных направлениях. На цепи 3’–5′ синтез дочерней полинуклеотидной цепи идет без перерывов; эта дочерняя цепь будет называться лидирующей. На цепи 5’–3′ — прерывисто, фрагментами (фрагменты Оказаки), которые после завершения репликации ДНК-лигазами сшиваются в одну цепь; эта дочерняя цепь будет называться запаздывающей (отстающей).
Особенностью ДНК-полимеразы является то, что она может начинать свою работу только с «затравки» (праймера). Роль «затравок» выполняют короткие последовательности РНК, образуемые при участи фермента РНК-праймазы и спаренные с матричной ДНК. РНК-затравки после окончания сборки полинуклеотидных цепочек удаляются.
Репликация протекает сходно у прокариот и эукариот. Скорость синтеза ДНК у прокариот на порядок выше (1000 нуклеотидов в секунду), чем у эукариот (100 нуклеотидов в секунду). Репликация начинается одновременно в нескольких участках молекулы ДНК. Фрагмент ДНК от одной точки начала репликации до другой образует единицу репликации — репликон.
Репликация происходит перед делением клетки. Благодаря этой способности ДНК осуществляется передача наследственной информации от материнской клетки дочерним.
Репарация («ремонт»)
Репарацией называется процесс устранения повреждений нуклеотидной последовательности ДНК. Осуществляется особыми ферментными системами клетки (ферменты репарации). В процессе восстановления структуры ДНК можно выделить следующие этапы: 1) ДНК-репарирующие нуклеазы распознают и удаляют поврежденный участок, в результате чего в цепи ДНК образуется брешь; 2) ДНК-полимераза заполняет эту брешь, копируя информацию со второй («хорошей») цепи; 3) ДНК-лигаза «сшивает» нуклеотиды, завершая репарацию.
Наиболее изучены три механизма репарации: 1) фоторепарация, 2) эксцизная, или дорепликативная, репарация, 3) пострепликативная репарация.
Изменения структуры ДНК происходят в клетке постоянно под действием реакционно-способных метаболитов, ультрафиолетового излучения, тяжелых металлов и их солей и др. Поэтому дефекты систем репарации повышают скорость мутационных процессов, являются причиной наследственных заболеваний (пигментная ксеродерма, прогерия и др.).
Строение и функции РНК
РНК — полимер, мономерами которой являются рибонуклеотиды. В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК). Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК.
Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов.
Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой.
Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.
Транспортные РНК содержат обычно 76 (от 75 до 95) нуклеотидов; молекулярная масса — 25 000–30 000. На долю тРНК приходится около 10% от общего содержания РНК в клетке. Функции тРНК: 1) транспорт аминокислот к месту синтеза белка, к рибосомам, 2) трансляционный посредник. В клетке встречается около 40 видов тРНК, каждый из них имеет характерную только для него последовательность нуклеотидов. Однако у всех тРНК имеется несколько внутримолекулярных комплементарных участков, из-за которых тРНК приобретают конформацию, напоминающую по форме лист клевера. У любой тРНК есть петля для контакта с рибосомой (1), антикодоновая петля (2), петля для контакта с ферментом (3), акцепторный стебель (4), антикодон (5). Аминокислота присоединяется к 3′-концу акцепторного стебля. Антикодон — три нуклеотида, «опознающие» кодон иРНК. Следует подчеркнуть, что конкретная тРНК может транспортировать строго определенную аминокислоту, соответствующую ее антикодону. Специфичность соединения аминокислоты и тРНК достигается благодаря свойствам фермента аминоацил-тРНК-синтетаза.
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК: 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК: 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы.
Строение и функции АТФ
Аденозинтрифосфорная кислота (АТФ) — универсальный источник и основной аккумулятор энергии в живых клетках. АТФ содержится во всех клетках растений и животных. Количество АТФ в среднем составляет 0,04% (от сырой массы клетки), наибольшее количество АТФ (0,2–0,5%) содержится в скелетных мышцах.
АТФ состоит из остатков: 1) азотистого основания (аденина), 2) моносахарида (рибозы), 3) трех фосфорных кислот. Поскольку АТФ содержит не один, а три остатка фосфорной кислоты, она относится к рибонуклеозидтрифосфатам.
Для большинства видов работ, происходящих в клетках, используется энергия гидролиза АТФ. При этом при отщеплении концевого остатка фосфорной кислоты АТФ переходит в АДФ (аденозиндифосфорную кислоту), при отщеплении второго остатка фосфорной кислоты — в АМФ (аденозинмонофосфорную кислоту). Выход свободной энергии при отщеплении как концевого, так и второго остатков фосфорной кислоты составляет по 30,6 кДж. Отщепление третьей фосфатной группы сопровождается выделением только 13,8 кДж. Связи между концевым и вторым, вторым и первым остатками фосфорной кислоты называются макроэргическими (высокоэнергетическими).
Запасы АТФ постоянно пополняются. В клетках всех организмов синтез АТФ происходит в процессе фосфорилирования, т.е. присоединения фосфорной кислоты к АДФ. Фосфорилирование происходит с разной интенсивностью при дыхании (митохондрии), гликолизе (цитоплазма), фотосинтезе (хлоропласты).
АТФ является основным связующим звеном между процессами, сопровождающимися выделением и накоплением энергии, и процессами, протекающими с затратами энергии. Кроме этого, АТФ наряду с другими рибонуклеозидтрифосфатами (ГТФ, ЦТФ, УТФ) является субстратом для синтеза РНК.
Источник