
Какие промежуточные продукты в цикле трикарбоновых кислот

Глюкоза как субстрат. Важнейшими и наиболее распространенными анаплеротическими реакциями, восполняющими расход промежуточных продуктов цикла трикарбоновых кислот у животных, растений и микроорганизмов, являются реакции карбоксилирования Сз-кислот (пирувата, фосфоенолпирувата), ведущие к образованию оксалоацетата [c.248]
Если во время роста клеток промежуточные продукты цикла трикарбоновых кислот используются в биосинтетических процессах, недостаток этих продуктов компенсируется восполняющими (анаплеротическими) реакциями. Одна из функций таких реакций-регенерация оксалоацетата, из которого в результате взаимодействия с ацетилкоферментом А образуется цитрат. [c.248]
Глюконеогенез —синтез глюкозы из неуглеводных продуктов. Такими продуктами или метаболитами являются в первую очередь молочная и пировиноградная кислоты, так называемые гликогенные аминокислоты, глицерол и ряд других соединений. Иными словами, предшественниками глюкозы в глюконеогенезе может быть пируват или любое соединение, превращающееся в процессе катаболизма в пируват или один из промежуточных продуктов цикла трикарбоновых кислот . [c.338]
Чаще всего водород отнимается от промежуточных продуктов цикла трикарбоновых кислот (ЦТК) – цикла Кребса (изолимонная, а-кетоглутаровая, янтарная и яблочная кислоты). Цикл Кребса – это за- [c.136]
Промежуточные продукты цикла трикарбоновых кислот [44] [c.198]
Катаболизм промежуточных продуктов цикла трикарбоновых кислот [c.326]
В ЭТОЙ главе рассматривается биосинтез аминокислот и некоторых молекул, которые из них образуются. Прежде всего мы рассмотрим реакции, приводящие к включению азота в состав аминокислот. Этот путь начинается с восстановления N2 до в клетках азотфиксирующих микроорганизмов. Затем NH4 включается в аминокислоты через глутамат и глутамин, два ключевых соединения азотистого метаболизма. Десять из основного набора двадцати аминокислот синтезируются из промежуточных продуктов цикла трикарбоновых кислот и других метаболических последовательностей с помощью несложных реакций. Мы рассмотрим эти биосинтетические пути и опишем биосинтез ароматических аминокислот и гистидина в качестве примеров аминокислот, синтезирующихся более сложным образом. На самом деле человек должен получать эти десять аминокислот с пищей, потому их и называют незаменимыми аминокислотами. В этих реакциях участвуют два весьма любопытных посредника тетрагидрофолят, многоцелевой переносчик одноуглеродных единиц трех степеней окисления, и 5-аденозилметионин, главный донор метильных групп. Еще одна важная сфера наших интересов-регуляция метаболизма аминокислот. На примере глу-тамин-синтетазы мы проиллюстрируем некоторые общие принципы регуляции. Конец настоящей главы посвящен синтезу и распаду гема. [c.230]
Эффективность данного процесса, можно повысить, изучив-механизмы, регуляции метаболизма в микрофлоре систем с активным илом. Регуляция биодеградации— это. сложная задача. Однако, зная биохимию соответствующих процессов, мы,. по-виДимому, сможем вмешиваться, и в их регуляцию. Например, добавление к илу промежуточных продуктов цикла трикарбоновых кислот в низ 11х концентрациях (2—5 мг/л), глюкозы, аминокислот и витаминов (в частности, аланина и никотиновой кислоты) приводит к ускорению окисления ряда соединений. Введение этих промежуточных продуктов в состав, биомассы увеличивает энергетические потребности системы, стимулирует синтез АТР за счет усиленного окисления неорганических веществ типа серы или аммиака. Понимание биохимии подобных процессов, видимо, даст возможность вмешиваться в процессы регуляции метаболизма. [c.255]
Отсутствие любого из промежуточных продуктов цикла трикарбоновых кислот должно остановить дальнейшее функционирование цикла, если только другой продукт цикла не будет регенерироваться со скоростью, равной скорости исчезновения первого субстрата. Чаще [c.120]
Как происходит пополнение оксалоацетата Млекопитающие лишены ферментативного механизма, обеспечивающего превращение ацетил-СоА в оксалоацетат или другой промежуточный продукт цикла трикарбоновых кислот. Оксалоацетат у них образуется путем карбоксилирования пирувата, катализируемого пируват-карбоксилазой (разд. 15.15). [c.64]
Пермеазы — переносчики специфических метаболитов через биомембраны. Они напоминают ферменты, так как обладают специфичностью по отношению к определенным веществам, способны насыщаться при высоких концентрациях субстрата и могут подвергаться специфическому ингибированию. В митохондриях печени крыс были идентифицированы переносчики для АДФ, АТФ, фосфата, а также для некоторых промежуточных продуктов цикла трикарбоновых кислот. Наиболее хорошо изучен переносчик внутренней мембраны, осуществляющий перенос АДФ и АТФ — соединений, которые не могут проникать через мембрану путем диффузии. [c.121]
Анаплеротнческие реакции — ферментативные реакции, пополняющие запас промежуточных продуктов цикла трикарбоновых кислот, их еще называют возмещающими. Наиболее важная реакция такого рода — ферментативное карбокси-лирование пирувата с образованием оксалоацетата, катализируемая пируваткарбок-силазой. Когда цикл трикарбоновых кислот функционирует в условиях недостатка оксалоацетата или других промежуточных продуктов, реакция карбоксилирования пирувата стимулируется, в результате чего запас оксалоацетата растет. При избытке оксалоацетата может происходить декарбоксилирование с образованием пирувата [c.96]
Расщепление органических соединений в процессе дыхания также сопровождается субстратным фосфорилированием, но основное количество АТФ образуется при окислительном фосфорилировании, сопряженном с окислением промежуточных продуктов цикла трикарбоновых кислот. Общий выход АТФ на одну молекулу субстрата, окисленного при дыхании, значительно выше, чем при его сбраживании. [c.47]
Таким образом, пируват-карбоксилаза не только имеет важное значение для глюконеогенеза, но играет также критическую роль в поддержании необходимой концентрации промежуточных продуктов цикла трикарбоновых кислот. Указанные промежуточные продукты должны все время восполняться, поскольку они потребляются при некоторых биосинтетических реакциях, например при синтезе гема. Эта роль пируват-карбоксилазы получила название анаплеротической, что означает восполняющая, компенсирующая. [c.108]
Аминокислоты синтезируются из промежуточных продуктов цикла трикарбоновых кислот и других важных метаболитов [c.233]
Рассматривая различные стороны возможной корреляции между липогенезом и синтезом полиеновых антибиотиков, Т. Т. Ефимова и др. (1977) предполагают, что их образование лимитируется количеством пропионил-КоА, так как среди многочисленных жирных кислот у изученных продуцентов практически полностью отсутствовали жирные кислоты нормального строения с нечетным числом атомов углерода в цепи, синтез которых начинается с пропионил-КоА. Основным источником этого продукта является метилмалонил-КоА, который, в свою очередь, происходит из сукционил-КоА — промежуточного продукта цикла трикарбоновых кислот. Усиленному образованию пропионил-КоА способствует, по мнению -авторов, низкая активность сукционатдегидрогеназы, что для A t. levoris показали результаты исследований О. С. Кузнецовой (1975). Не исключается значение и других путей образования пропионил-КоА. [c.176]
Пируваткарбоксилаза-это митохондриальный фермент. Образовавшийся С-ок-салоацетат смешивается с пулом оксалоацетата, используемого в цикле трикарбоновых кислот. Следовательно, между С-оксалоацетатом и промежуточными продуктами цикла трикарбоновых кислот устанавливается равновесие с образованием через С-сукцинат смеси 1- С- и 4- С-оксалоацетата. Из оксалоацетата, меченного С в положении 1, образуется 3,4- С-глюкоза [см. задачу 2(6)]. [c.722]
Цикл трикарбоновых кислот не только выполняет функцию конечного окисления органических веществ, но и обеспечивает процессы биосинтеза различными предшественниками, такими как 2-оксоглутарат, оксалоацетат и сукцинат. Отсутствие этих кислот привело бы к нехватке оксалоацетата, который служит акцептором для ацетил-СоА, и тем самым к нарушению цикла. Восполнение потерь промежуточных продуктов цикла трикарбоновых кислот-функция так называемых анапле-ротических реакций. Важнейший механизм обеспечения цикла С4-дикарбоновыми кислотами состоит в карбоксилировании пирувата и фосфоенолпирувата (С3 + С – С ). Эти реакции будут подробнее рассмотрены позже (разд. 7.5). [c.234]
В качестве субстратов окисления (т. е. веществ, от которых отнимается водород) в тканевом дыхании используются разнообразные промежуточные продукты распада белков, углеводов и жиров. Однако наиболее часто окислению подвергаются промежуточные продукты цикла трикарбоновых кислот (ЦТК) – цикла Кребса (изолимонная, а-кетоглутароаая, янтарная и яблочная кислоты). Цикл Кребса – это за-верщающий этап катаболизма, в ходе которого происходит окисление [c.37]
До сих пор, обсуждая цикл трикарбоновых кислот, мы рассматриваем его как основной путь расщепления, обеспечивающий генерирование АТР. Однако цикл трикарбоновых кислот выполняет и другую роль он поставляет промежуточные продукты для процессов биосинтеза (рис. 13.15). Например, большинство углеродных атомов в порфи-ринах происходит из сукцинил-СоА, Многие аминокислоты происходят из а-оксоглута-рата и оксалоацетата. Биосинтез этих соединений мы рассмотрим в последующих главах. Здесь же необходимо отметить тот важный момент, что потребление промежуточных продуктов цикла трикарбоновых кислот для целей биосинтеза обязательно должно сопровождаться их пополнением. Предположим, например, что оксалоацетат превращается в аминокислоты, используемые для биосинтеза белка. Если при этом не будет происходить синтеза оксалоацетата de novo, то функционирование цикла трикарбоновых кислот прекратится, поскольку ацетил-СоА может включиться в цикл только после конденсации с оксалоацетатом. [c.64]
Активность пируват-карбоксилазы зависит от присутствия ацетил-СоА. В отсутствие связанного с ферментом аиетил-СоА (или другого близкого к нему ацил-СоА) биотин не карбоксилируется. Вторая частичная реакция не зависит от ацетил-СоА. Аллостерическая активация пируват-карбоксилазы при участии ацетил-СоА представляет собой важный физиологический механизм контроля. Оксалоацетат, продукт пируват-карбоксилазной реакции, является одновременно и стехиометрическим промежуточным продуктом глюконеогенеза, и каталитическим промежуточным продуктом цикла трикарбоновых кислот. Высокое содержание ацетил-СоА служит сигналом необходимости большего количества оксалоацетата. Если имеет место избыток АТР, оксалоацетат потребляется в процессе глюконеогенеза. В условиях недостатка АТР оксалоацетат включается в цикл трикарбоновых кислот, конденсируясь с аце-тил-СоА. [c.108]
Аспартат, аминокислота, содержащая четыре атома углерода, прямо трансаминирует-ся в оксалоацетат, промежуточный продукт цикла трикарбоновых кислот [c.168]
Глутамат синтезируется из NH4 и а-оксо-глутарата, промежуточного продукта цикла трикарбоновых кислот, под действием глутамат-дегидрогеназы. Мы уже встречались с этим ферментом в разделе, посвященном распаду аминокислот (разд. 18.1). Когда реакция идет в направлении биосинтеза, используется NADPH в качестве восстановителя когда же реакция является катаболиче-ской, в ней участвует NAD в качестве [c.232]
Биохимия Т.3 Изд.2 (1985) — [
c.64
,
c.65
]
Источник
История изучения
Биологическая роль некоторых реакций цикла Кребса (ЦК) была изучена американским биохимиком венгерского происхождения Альбертом Сент-Дьердьи. В частности, он выделил ключевой компонент ЦТК — фумарат. Исследования в этом направлении продолжил Ганс Кребс. В итоге он установил всю последовательность реакций и соединений, образующиеся на всех этапах процесса. Ученый не смог определить, с преобразования какой кислоты начинается цикл — лимонной или изолимонной. Сейчас известно, что это лимонная кислота. Поэтому ЦК называют также цитратным или циклом лимонной кислоты.
Позднее американец Альберт Ленинджер, занимающийся биоэнергетикой, определил, что все реакции ЦК протекают в митохондриях клеток. С получением доступа к изотопам углерода появилась возможность более досконального изучения и уточнения данных о промежуточных соединениях на разных этапах цикла.
С пищей в организм поступают три основные группы сложных биохимических соединений — белки, жиры и углеводы. Они являются первичными метаболитами, потому что участвуют в обмене веществ или в метаболизме. Этот процесс происходит между любыми живыми клетками и окружающей средой непрерывно. Суть цикла Кребса заключается в том, что он является областью схождения двух путей метаболизма. Это следующие процессы:
- катаболизм, при котором происходит распад более сложных веществ на простые, в частности, глюкозы на моносахариды;
- анаболизм — синтез сложных веществ из простых, например, белков из аминокислот.
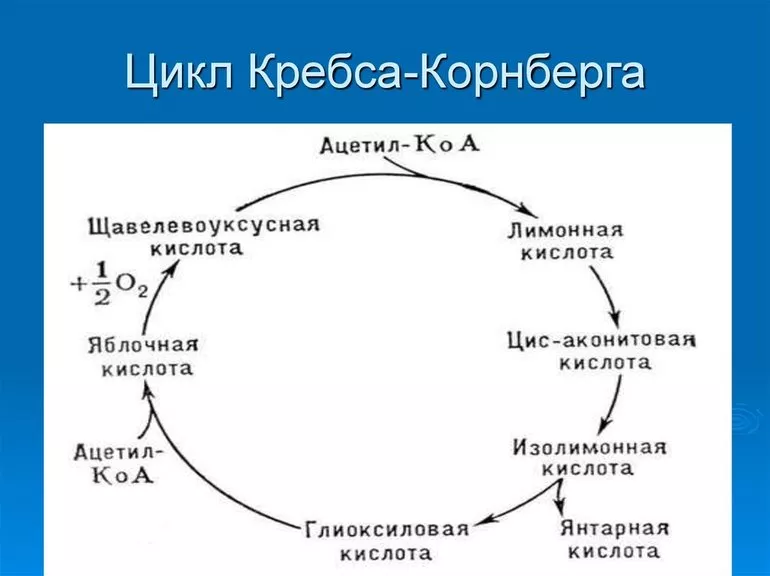
После попадания в пищеварительную систему сложные вещества расщепляются под действием ферментов на более простые, которые внутри клеток превращаются сначала в пируват (пировиноградную кислоту), а затем — в ацетильный остаток. Все эти преобразования можно назвать подготовкой к ЦК, а образование остатка — его запуском или начальным этапом.
Дальнейшие стадии цикла трикарбоновых кислот являются частью катаболизма. Процесс идет каскадно. Каждый предыдущий этап запускает последующий, а промежуточные продукты химических реакций служат не только для продолжения цикла, но и при определенных потребностях организма могут пополнять запасы веществ, необходимых для синтеза новых соединений (анаболизма).
Клеточное дыхание
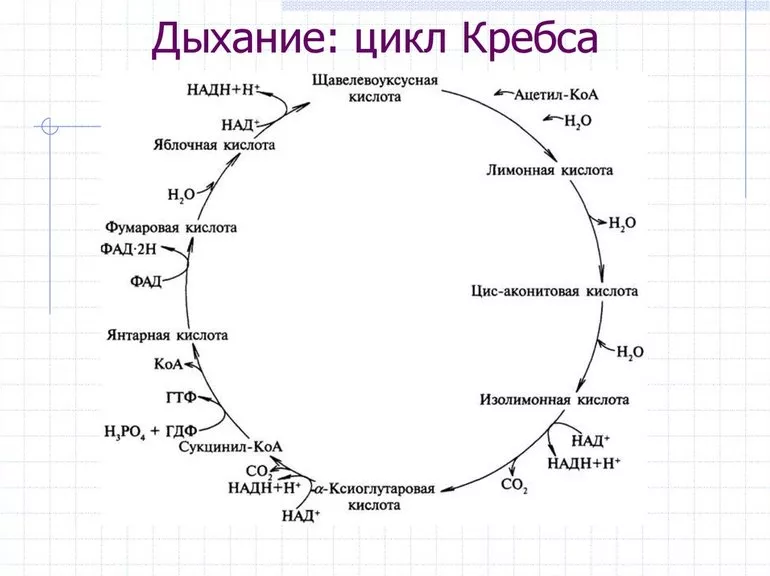
Для нормальной жизнедеятельности живым клеткам постоянно требуется энергия. Ее главный универсальный источник — аденозинтрифосфат (АТФ), способный встраиваться в белки организма напрямую. Это соединение получается в результате ряда реакций окисления, носящих общее название «клеточное дыхание». При этом происходит постепенный распад органических веществ вплоть до простейших неорганических — углекислого газа CO2 и воды H2O.
Структурное строение молекул АТФ содержит фосфорангидридные связи, которые имеют свойство накапливать высвобожденную при прохождении реакций клеточного дыхания энергию, поэтому называются макроэргическими. Так создаются энергетические запасы клеток, которые могут высвобождается при необходимости разрывом этих связей. Процесс синтеза АТФ и класса вспомогательных соединений включает три этапа:
- Гликолиз происходит в цитоплазме.
- В матриксе митохондрий проходят все химические реакции цикла Кребса.
- Окислительное фосфорилирование на внутренней мембране митохондрий.
Преобразование аденозиндифосфата (АДФ) в АТФ характерно для всех этапов. Но наибольшее суммарное количество молекул с макроэргическими связями образуется при фосфорилировании. Это не значит, что процессы гликолиза и ЦК менее важны. Многие соединения, образующиеся во время их протекания, участвуют в регуляции клеточного дыхания.
Описание процесса
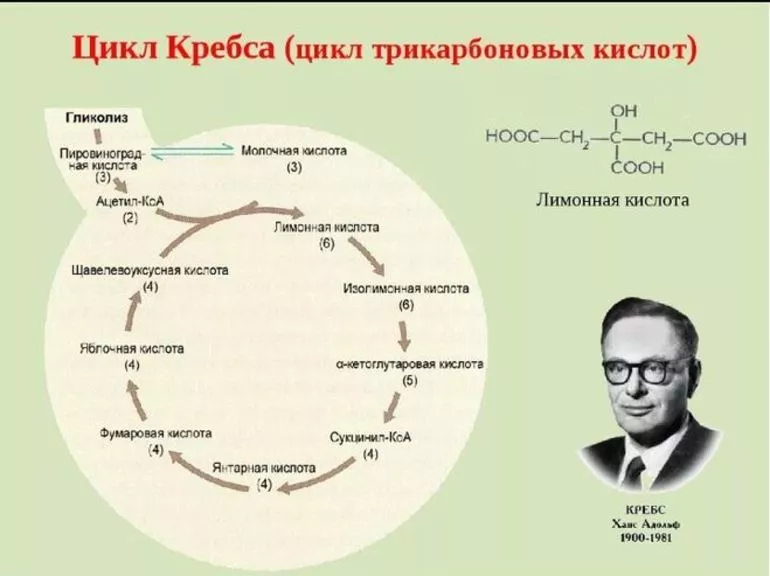
Протекание ЦК достаточно экономно с точки зрения энергозатрат. Такой эффект достигается благодаря тому, что он связывает два метаболических направления. В процесс вовлекаются вещества, подлежащие утилизации, которые либо служат энергетическим «топливом», либо возвращаются в круг анаболизма. Подготовительная стадия ЦК заключается в распаде глюкозы, аминокислот и жирных кислот на молекулы пирувата или лактата.
Органеллы митохондрий способны преобразовывать пируват в ацетильный остаток (ацетил-коэнзим А или ацетил-КоА), представляющий собой вместе с тиольной группой, которая может его переносить, кофермент А. Некоторое соединения могут сразу распадаться до ацетил-КоА, минуя стадию пирувата. При этом пировиноградная кислота может вовлекаться непосредственно в ЦК, не преобразуясь в ацетил-КоА.
Начальные этапы
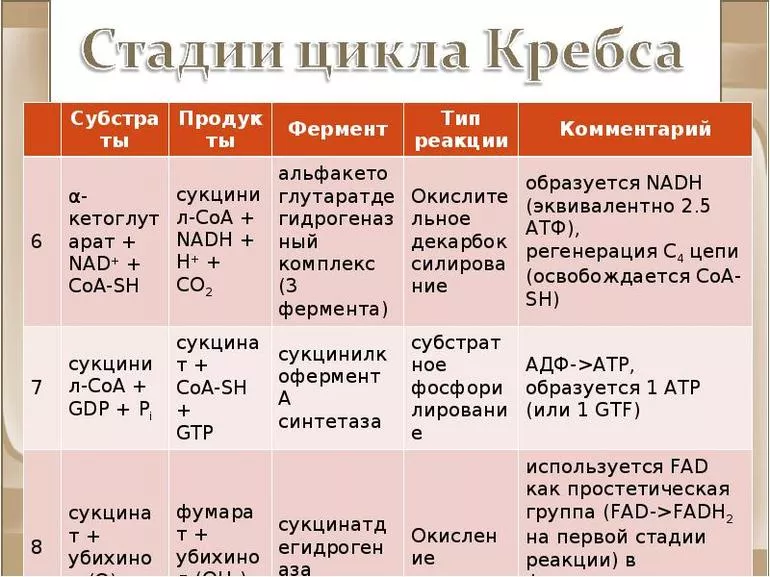
Первая стадия необратима и состоит из конденсации ацетил-КоА с четырехуглеродным веществом — оксалоацетатом (щавелевоуксусной кислотой или ЩУК), что приводит к образованию шестиуглеродного цитрата (лимонной кислоты). Во время реакции метильная группа ацетил-КоА соединяется с карбонильной группой ЩУК. Благодаря быстрому гидролизу промежуточного соединения цитроил-КоА этот этап проходит без затрат энергии извне.
На второй стадии образуется изоцитрат (изолимонная кислота) из цитрата через цис-аконитат. Это реакция обратимой изомеризации через образование промежуточной трикарбоновой кислоты, в которой катализатором выступает фермент аконитатгидратаза.
Далее происходит дегидрирование и декарбоксилирование изоцитрата до промежуточного соединения оксалосукцинат с выделением углекислого газа. После декарбоксилирования оксалосукцината образуется енольное соединение, которое перестраивается и превращается в пятиуглеродную кислоту — α-кетоглутарат (оксоглутарата), чем и завершает третью ступень ЦК. Четвертый этап — α-кетоглутарат декарбоксилирует и реагирует с ацетил-КоА. При этом получается сукцинил-КоА, соединение янтарной кислоты и коэнзима-А, выделяется СО2.
Замыкание цикла
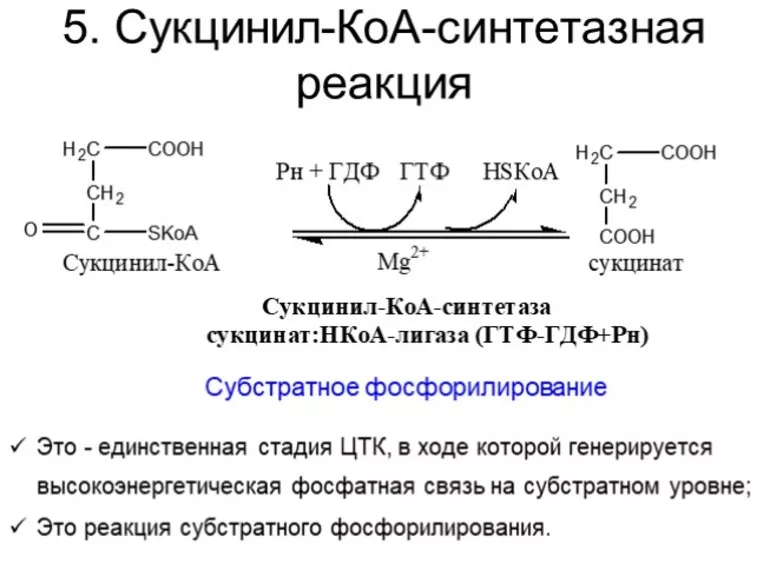
На пятой стадии сукцинил-КоА преобразуется в сукцинат (янтарную кислоту). Для этого этапа характерно субстратное фосфолирование, подобное синтезу АТФ при гликолизе. Введение в ЦК фосфорной группы РО3 становится возможным благодаря присутствию фермента ГДФ (гуанозиндифосфата) или АДФ (аденозиндифосфата), которые в процессе синтеза сукцината из дифосфатов становятся трифосфатами.
Начиная с шестой стадии, цикл начинает постепенно замыкаться. Сначала сукцинат под действием каталитического фермента сукцинатдегидрогеназы дегидрирует до фумарата. Дальнейшее дигидрирование приводит к седьмому этапу — образованию L-малата (яблочной кислоты) из фуратата через переходное соединение с карбанионом.
Последняя реакция цикла трикарбоновых кислот малат окисляется до щавелевоуксусной кислоты. Первая стадия следующего ЦК начинается с новой молекулы ацетил-КоА.
Значение и функции
Этот восьмиэтапный циклический процесс, итогом которого является окисление ацетильного остатка до углекислого газа, может показаться излишне сложным. Тем не менее, он имеет огромное значение в метаболизме промежуточных реакций и выполняет ряд функций. К ним относятся:
- энергетическая;
- анаболическая;
- катаболическая;
- транспортная.
Цикл Кребса участвуют в катаболизме жиров и углеводов. Соединения, образующиеся на разных стадиях процесса, участвуют в синтезе многих необходимых для организма веществ — глутамина, порфиринов, глицина, фенилаланина, цистеина и других. Когда промежуточные продукты покидают ЦК для участия в синтезе, происходит их замещение с помощью так называемых анаплеротических реакций, которые катализируются регуляторными ферментами, например, пируваткарбоксилазой.
Транспортная функция ЦК заключается в содействии гликолизу. Глюкозу невозможно превратить сразу в АТФ, поэтому механизм гликолиза действует поэтапно и сопровождается постоянным перемещением атомов и катионов водорода от одних соединений к другим. Для их транспортировки нужны специальные соединения, которые получаются на одной из стадий ЦТК. Участвующие в гликолизе коферменты цикла Кребса:
- НАД*H+(Никотинамидадениндинуклеотид с катионом водорода). Образуется на III стадии ЦК.
- ФАД*H2 (Флавинадениндинуклеотид с молекулой водорода). Появляется на V стадии ЦК.
Реакции ЦК имеют и большое клиническое значение. Хотя для людей не свойственны мутации, связанные с генами ферментов, участвующих в цикле, однако их редкие проявления губительны для здоровья. Они могут приводить к опухолям мышц и почек, нарушениям работы нервной системы.
Существует множество видов визуального и слухового отображения цикла Кребса — схемы с формулами, уравнения химических реакций, разнообразные таблицы и даже мнемонические способы для полного запоминания его главных «участников».
Источник