
Какие продукты обмена они выделяются

Обмен веществ в организме. Пластическая и энергетическая роль питательных веществ
Постоянный обмен веществ и энергии между организмом и окружающей средой является необходимым условием его существования и отражает их единство. Сущность этого обмена заключается в том, что поступающие в организм питательные вещества после пищеварительных превращений используются как пластический материал. Энергия, образующаяся при этих превращениях восполняет энергозатраты организма.
Синтез сложных специфичных веществ организма из простых соединений, всасывающихся в кровь из пищеварительного канала, называется ассимиляцией или анаболизмом. Распад веществ организма до конечных продуктов, сопровождающийся выделением энергии называется диссимиляцией или катаболизмом. Два этих процесса неразрывно связаны. Ассимиляция обеспечивает аккумуляцию энергии, а энергия выделяющаяся при диссимиляции необходима для синтеза веществ. Анаболизм и катаболизм объединены в единый процесс с помощью АТФ и НАДФ. С их помощью энергия образующаяся в результате диссимиляции передается для процессов ассимиляции.
Белки в основном являются пластическим материалом. Они входят в состав клеточных мембран, органелл. Белковые молекулы постоянно обновляются. Но это обновление происходит не только за счет белков пищи, но и посредством реутилизации собственных белков организма. Из 20 аминокислот, образующих белки 10 являются незаменимыми. Т.е. не могут образовываться в организме. Конечными продуктами распада белков являются такие азотсодержащие соединения, как мочевина, мочевая кислота, креатинин.
Состояние белкового обмена оценивается по азотистому балансу. Это соотношение количества азота поступающего с белками пищи и выделенного из организма с азотсодержащими продуктами обмена. В белке содержится около 16 г азота. Следовательно выделение 1 г азота свидетельствует о распаде в организме 6,25 г белка. Если количество выделяемого азота равно количеству поглощенного организмом имеет место азотистое равновесие. Если поступившего азота больше, чем выделенного, это называется положительным азотистым балансом. В организме происходит задержка или ретенция азота. Положительный азотистый баланс наблюдается при росте организма, при выздоровлении после тяжелых заболевания, сопровождавшихся похуданием и после длительного голодания. Когда количество азота, выделенного организмом больше, чем поступившего, имеет место отрицательный азотистый баланс. Его возникновение объясняется распадом собственных белков организма. Он возникает при голодании, отсутствии в пище незаменимых аминокислот, нарушениях переваривания и всасывания белка, тяжелых заболеваниях. Количество белка которое полностью обеспечивает потребности организма называется белковым оптимумом. Минимальное, обеспечивающее лишь сохранение азотистого баланса – белковым минимумом. ВОЗ рекомендует потребление белка не менее 0,75 г на кг веса в сутки. Энергетическая роль белков относительно небольшая.
Жирами организма являются триглицериды, фосфолипиды и стерины. Они также имеют определенную пластическую роль, так как фосфолипиды, холестерин, жирные кислоты входят в состав клеточных мембран и органелл. Основная их роль энергетическая. При окислении липидов выделяется наибольшее количество энергии, поэтому около половины энергозатрат организма обеспечивается липидами. Кроме того, они являются аккумулятором энергии в организме потому что откладываются в жировых депо и используются по мере необходимости. Жир депо составляют около 15% веса тела. Покрывая внутренние органы жировая ткань выполняет и пластическую функцию. Например околопочечный жир способствует фиксации почек и предохранению их от механических воздействий. Липиды являются источниками воды, потому что при окислении 100 г жира образуется около 100 г воды. Особую функцию выполняет бурый жир, располагающийся вдоль крупных сосудов. Содержащийся в его жировых клетках полипептид тормозит ресинтез АТФ за счет липидов. В результате резко усиливается теплопродукция. Большое значение имеют незаменимые жирные кислоты – линолевая, линоленовая и арахидоновая. Они не образуются в организме. Без них невозможен синтез фосфолипидов клеток, образование простагландинов и т.д. При их отсутствии задерживается рост и развитие организма.
Углеводы в основном играют энергетическую роль, т.к. служат основным источником энергии для клеток. Потребности нейронов покрываются исключительно глюкозой. Углеводы аккумулируются в виде гликогена в печени и мышцах. Углеводы имеют определенное пластическое значение. Глюкоза необходима для образования нуклеотидов и синтеза некоторых аминокислот.
Методы измерения энергетического баланса организма
Соотношение между количеством энергии, поступившей в организм с пищей, и энергии, выделенной организмом во внешнюю среду называется энергетическим балансом организма. Существует 2 метода определения выделяемой организмом энергии.
- Прямая калориметрия. Принцип прямой калориметрии основан на том, что все виды энергии в конечном итоге переходят в тепловую. Поэтому при прямой калориметрии определяют количество тепла выделяемого организмом в окружающую среду за единицу времени. Для этого используют специальные камеры с хорошей теплоизоляцией и системой теплоообменных труб, в которых циркулирует и нагревается вода.
- Непрямая калориметрия. Она заключается в определении соотношения выделенного углекислого газа и поглощенного кислорода за единицу времени. Т.е. полном газовом анализе. Это соотношение называется дыхательным коэффициентом (ДК).
Величина дыхательного коэффициента определяется тем, какое вещество окисляется в клетках организма. Например в молекуле углеводов атомов кислорода много, Поэтому на их окисление кислорода идет меньше и их дыхательный коэффициент равен 1. В молекуле липидов кислорода значительно меньше, поэтому дыхательный коэффициент при их окислении 0,7. Дыхательный коэффициент белков 0,8. При смешанном питании его величина 0,85-0,9. Дыхательный коэффициент становится больше 1 при тяжелой физической работе, ацидозе, гипервентиляции и преобразовании в организме углеводов в жиры. Меньше 0,7 он бывает при переходе жиров в углеводы. Исходя из дыхательного коэффициента рассчитывается калорический эквивалент кислорода, т.е. количество энергии выделяемой организмом при потреблении 1 л кислорода. Его величина также зависит от характера окисляемых веществ. Для углеводов он составляет 5 ккал, белков 4,5 ккал, жиров 4,7 ккал. Непрямая калориметрия в клинике производится с помощью аппаратов “Метатест-2”, “Спиролит”.
Величина поступившей в организм энергии определяется количеством и энергетической ценностью пищевых веществ. Их энергетическую ценность определяют путем сжигания в бомбе Бертло в атмосфере чистого кислорода. Таким путем получают физический калорический коэффициент. Для белков он равен 5,8 ккал/г, углеводов 4,1 ккал/г, жиров 9,3 ккал/г. Для расчетов используют физиологический калорический коэффициент. Для углеводов и жиров он соответствует физическому, а для белков составляет 4,1 ккал/г. Его меньшая величина для белков объясняется тем, что в организме они расщепляются не до углекислого газа и воды, а да азотсодержащих продуктов.
Основной обмен
Количество энергии, которое затрачивается организмом на выполнение жизненно важных функций называется основным обменом. Это затраты энергии на поддержание постоянства температуры тела, работу внутренних органов, нервной системы, желез. Основной обмен измеряется методами прямой и непрямой калориметрии при базисных условиях, т.е. лежа с расслабленными мышцами, при температуре комфорта, натощак. Согласно закону поверхности, сформулированному в 19 веке Рубнером и Рише, величина основного прямопропорциональна площади поверхности тела. Это связано с тем, что наибольшее количество энергии тратится на поддержание постоянства температуры тела. Помимо этого на величину основного обмена влияют пол, возраст, условия окружающей среды, характер питания, состояние желез внутренней секреции, нервной системы. У мужчин основной обмен на 10% больше, чем у женщин. У детей его величина относительно веса тела больше, чем в зрелом возрасте, а у пожилых наоборот меньше. В холодном климате или зимой он возрастает, летом снижается. При гипертиреозе он значительно увеличивается, а гипотиреозе снижается. В среднем величина основного обмена у мужчин 1700 ккал/сут., а у женщин 1550.
Общий обмен энергии
Общий обмен энергии это сумма основного обмена, рабочей прибавки и энергии специфически-динамического действия пищи. Рабочая прибавка это энергетические затраты на физическую и умственную работу. По характеру производственной деятельности и энергозатратам выделяют следующие группы работающих:
- Лица умственного труда (преподаватели, студенты, врачи и т.д.). Их энергозатраты 2200-3300 ккал/сут.
- Работники занятые механизированным трудом (сборщики на конвейере). 2350-3500 ккал/сут.
- Лица занятые частично механизированным трудом (шофера). 2500-3700 ккал/сут.
- Занятые тяжелым немеханизированным трудом (грузчики). 2900-4200 ккал/сут. Специфически-динамическое действие пищи это энергозатраты на усвоение питательных веществ. Наиболее выражено это действие у белков, меньше у жиров и углеводов. В частности белки повышают энергетический обмен на 30%, а жиры и углеводы на 15%.
Физиологические основы питания. Режимы питания
В зависимости от возраста, пола, профессии потребление белков, жиров и углеводов должно составлять: у мужчин I-IV групп Б: 96-108 г, Ж: 90-120 г, У: 382-552 г; у женщин I-IV групп Б: 82-92 г, Ж: 77-102 г, У: 303-444 г.
В прошлом веке Рубнер сформулировал закон изодинамии, согласно которому пищевые вещества могут взаимозаменяться по своей энергетической ценности. Однако он имеет относительное значение, так как белки, выполняющие пластическую роль, не могут синтезироваться из других веществ. Это же касается незаменимых жирных кислот. Поэтому требуется питание сбалансированное по всем питательным веществам. Кроме того необходимо учитывать усвояемость пищи. Это соотношение всосавшихся и выделившихся с калом питательных веществ. Наиболее легко усваиваются животные продукты. Поэтому животный белок должен составлять не менее 50% суточного белкового рациона, а жиры не более 70% жирового.
Под режимом питания подразумевается кратность приема пищи и распределение ее калорийности на каждый прием. При трехразовом питании на завтрак должно приходится 30% калорийности суточного рациона, обед 50%, ужин 20%. При более физиологичном четырехразовом, на завтрак 30%, обед 40%, полдник 10%, ужин 20%. Интервал между завтраком и обедом не более 5 часов, а ужин должен быть не менее чем за 3 часа до сна. Часы приема пищи должны быть постоянными.
Обмен воды и минеральных веществ
Содержание воды в организме в среднем 73%. Водный баланс организма поддерживается путем равенства потребляемой и выделяемой воды. Суточная потребность в воде составляет 20-40 мл/кг веса. С жидкостями поступает около 1200 мл воды, пищей 900 мл и 300 мл образуется в процессе окисления питательных веществ. Минимальная потребность в воде составляет 1700 мл. При недостатке воды наступает дегидратация и если ее количество в организме снижается на 20% наступает смерть. Избыток воды сопровождается водной интоксикацией с возбуждением ЦНС и судорогами.
Натрий, калий, кальций, хлор необходимы для нормального функционирования всех клеток, в частности обеспечения механизмов формирования мембранного потенциала и потенциалов действия. Суточная потребность в натрии и калии 2-3 г, кальции 0,8 г, хлоре 3-5 г. Большое количество кальция находится в костях. Кроме того он нужен для свертывания крови, регуляции клеточного метаболизма. Основная масса фосфора также сосредоточена в костях. Одновременно входит а состав фосфолипидов мембран, участвует в процессах метаболизма. Суточная потребность в нем 0,8 г. Большая часть железа содержится в гемоглобине и миоглобине. Оно обеспечивает связывание кислорода. Фтор входит в состав эмали зубов. Сера в состав белков и витаминов. Цинк является компонентом ряда ферментов. Кобальт и медь необходимы для эритропоэза. Потребность во всех этих микроэлементах от десятков до сотен мг в сутки.
Регуляция обмена веществ и энергии
Высшие нервные центры регуляции энергетического обмена и обмена веществ находятся в гипоталамусе. Они влияют на эти процессы через вегетативную нервную систему и гипоталамо-гипофизарную систему. Симпатический отдел ВНС стимулирует процессы диссимиляции, парасимпатический ассимиляцию. В нем же находятся центры регуляции водно-солевого обмена. Но главная роль в регуляции этих базисных процессов принадлежит железам внутренней секреции. В частности инсулин и глюкагон регулируют углеводный и жировой обмены. Причем инсулин тормозит выход жира из депо. Глюкокортикоиды надпочечников стимулируют распад белков. Соматотропин наоборот усиливает синтез белка. Минералокортикоиды натрий-калиевый. Основная роль в регуляции энергетического обмена принадлежит тиреоидным гормонам. Они резко усиливают его. Они же главные регуляторы белкового обмена. Значительно повышает энергетический обмен и адреналин. Большое его количество выделяется при голодании.
Источник
Образовавшиеся при распаде пищи конечные продукты метаболизма либо выводятся через покровы тела и стенки трахей (CO2), либо абсорбируются в задней кишке (H2O), либо удаляются с остатками непереваренной пищи — экскрементами (мочевина, мочевая кислота, аммиак и др.).
При гидролизе нуклеиновых кислот образуются углеводы, фосфорная кислота и обогащённые азотом пуриновые (аденин, гуанин) или пиримидиновые (цитозин, тимин) основания. В свою очередь, пуриновые основания, подвергаясь окислению и дезаминированию, дают начало мочевой кислоте и её производным: аллантоину, аллантоиновой кислоте, мочевине и аммиаку, которые выводятся из организма. Пиримидиновые основания, хотя и способны преобразовываться в мочевину и аммиак, обычно вновь вовлекаются в метаболические процессы.
При гидролизе белков образуются аминокислоты и некоторые из них — чаще всего богатые азотом аргинин и гистидин — входят в состав экскрементов (в весьма малых количествах). Обычно они используются в синтезе пуриновых оснований, образуя наряду с ними мочевину. Таким образом, конечные продукты метаболизма азотсодержащих соединений формируются при окислении пуринов или синтезируются из аминокислот (рис. 100).
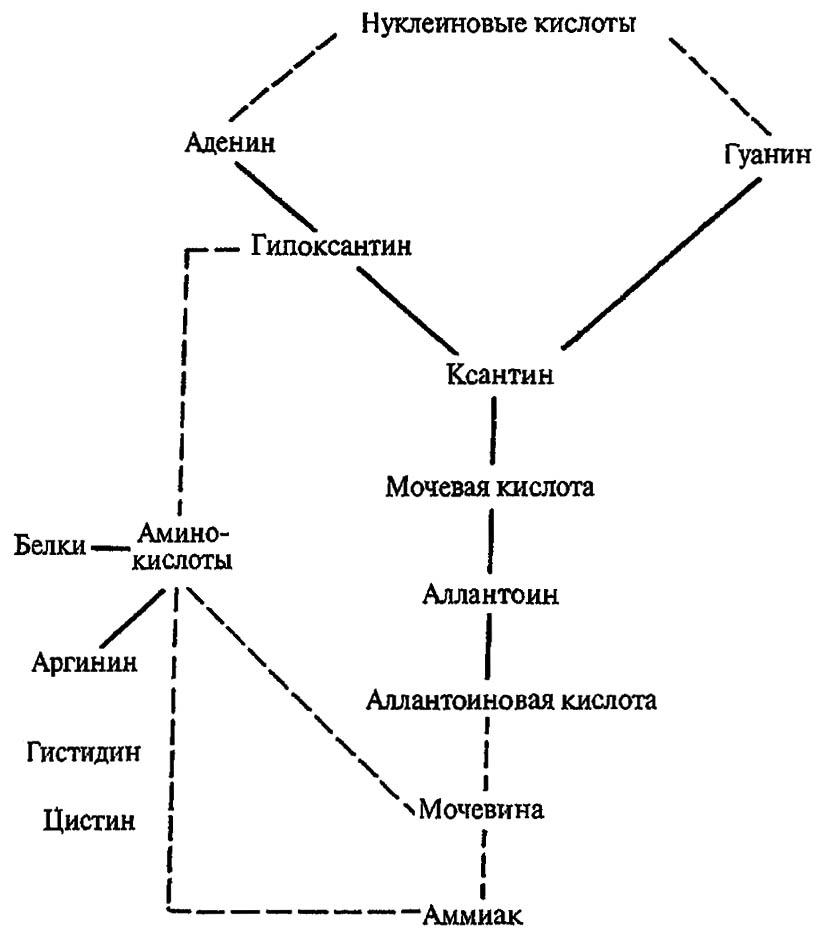
Рисунок 100. Конечные продукты обмена азотсодержащих соединений и их превращения у насекомых (по Gillot, 1980)
Большинство наземных насекомых выделяют азот в виде слаборастворимых и нетоксичных для организма мочевой кислоты, аллантоина и аллантоиновой кислоты. Они удаляются вместе с обезвоженными экскрементами; при этом возможные потери влаги сводятся к минимуму. Водорастворимые и токсичные даже в малых концентрациях мочевина и аммиак требуют для выведения очень больших количеств воды. Не случайно, что именно эти соединения являются конечными продуктами метаболизма у водных форм. Прежде чем поступить в заднюю кишку, в формирующиеся здесь экскременты, все эти метаболиты накапливаются в гемолимфе и извлекаются из неё специализированными органами выделения — мальпигиевыми сосудами.
Мальпигиевы сосуды представляют собой длинные и тонкие трубочки, впадающие в кишечник на уровне пилорического отдела (см. рис. 81). Вместе с задней кишкой они обеспечивают экскрецию азотсодержащих метаболитов и постоянство ионного баланса гемолимфы. Лишь у ногохвосток, некоторых двухвосток и тлей они не развиты.
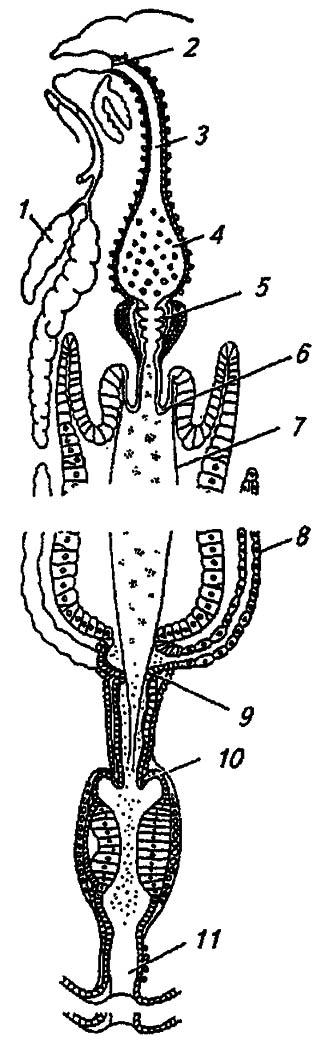
Рисунок 81. Схема кишечного тракта насекомых (по Шванвичу, 1949):
1 — слюнные железы; 2 — глотка; 3 — пищевод; 4 — зоб; 5 — провентрикулус; 6 — кардиальный клапан; 7 — перитрофическая мембрана; 8 — мальпигиев сосуд; 9, 10 — соответственно пилорический и ректальный клапаны; 11 — анус
Стенки сосудов образованы однослойным эпителием и мышечными волокнами. Оплетённые трахеями, но лишённые нервов, они способны только к миогенным червеобразным движениям. У щетино-хвосток, уховёрток и трипсов мальпигиевы сосуды не имеют мышц и пассивно колеблются в токах гемолимфы.
В простейшем случае, например у прямокрылых, мальпигиевы сосуды однообразны по всей длине и лишь насасывают плазму с содержащимися в ней экскретами (рис. 101). Далее эта «первичная моча» проникает в полость задней кишки и подвергается здесь реабсорбции. Все метаболически ценные вещества (H2O, Cl-, Na+, K+ и др.) возвращаются в гемолимфу, а экскреты выводятся из организма. Сравнительно малая эффективность работы таких сосудов компенсируется их громадным числом (до 250 и более).
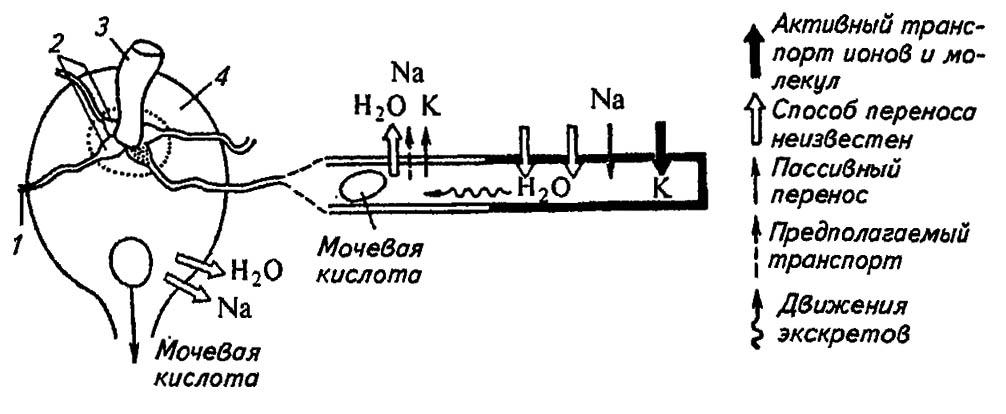
Рисунок 101. Строение и принципы работы мальпигиевых сосудов палочника (по Тыщенко, 1976):
1 — мальпигиевы сосуды; 2 — ампула; 3 — средняя кишка; 4 — задняя кишка
Сходным образом функционируют малочисленные (4–8) мальпигиевы сосуды некоторых жуков, однако их свободные концы врастают в стенку задней кишки. Высасывая из её полости воду, они энергично проводят первичную мочу, но не способны к её реабсорбции. У многих клопов происходит дифференциация отделов и эпителия сосудов и соответственно распределение функций по их длине. В дистальном отделе эпителиальные клетки несут плотный рабдориум и содействуют образованию первичной мочи. Переходя в проксимальный отдел, клетки которого снабжены рыхлым рабдориумом, она подвергается реабсорбции, и, таким образом, этот отдел принимает на себя функции задней кишки прямокрылых (рис. 102).
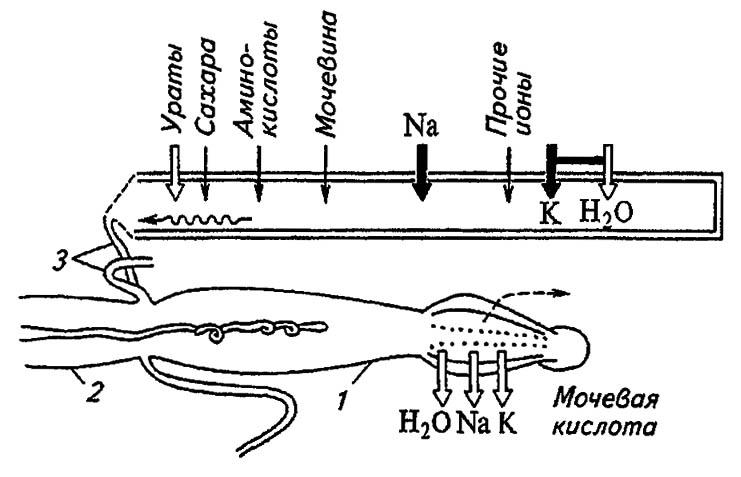
Рисунок 102. Строение и принципы работы мальпигиевых сосудов клопа Rhodnius prolixus St. (по Тыщенко, 1976):
1 — задняя кишка; 2 — средняя кишка; 3 — мальпигиевы сосуды
Ещё большей сложностью строения отличаются мальпигиевы сосуды двукрылых. Наряду с дистальным и проксимальным отделами в них выделяются промежуточный и медиальный отделы. В дистальном происходит насасывание мочевой кислоты и её солей, а также ионов Ca2+, тогда как в промежуточном и медиальном — воды. В проксимальком отделе реабсорбируются метаболически ценные продукты. У гусениц многих бабочек свойства сосудов, отмеченные у клопов и двукрылых, сочетаются с криптонефрией (рис. 103).
Рисунок 103. Строение и принципы работы мальпигиевых сосудов гусеницы бабочки Corcyra cephalonica (по Тыщенко, 1976):
1 — средняя кишка; 2 — тонкая кишка; 3 — ампула мальпигиева сосуда; 4 — прямая кишка
Заполняющая мальпигиевы сосуды жидкость изотонична гемолимфе, но отличается от неё по набору ионов. В частности, у палочника Carausius morosus Вr. ионы K+ преобладают внутри сосуда, а ионы Na+ — снаружи. Нарушение ионного баланса проявляется в разности потенциалов и возникновении электрохимического градиента.
Ионы K+ активно транспортируются внутрь и, по-видимому, переносят молекулы воды вопреки градиенту диффузии. Несколько по-иному работают мальпигиевы сосуды кровососущего клопа Rhodnius prolixus St. В них активно проникают ионы K+ и Na+, транспортирующие воду. Экскреты, поступающие в ихдистальные отделы в виде мочекислых солей натрия и калия, оказываются в слабощелочной среде (рН 7,2), но, продвигаясь проксимально, встречают слабокислую реакцию (рН 6,6) жидкости. В этих условиях Na+ и K+ освобождаются, а мочевая кислота кристаллизуется и выпадает в осадок (см. рис. 102).
Активность экскреции у Rhodnius prolixus St. существенно повышается (в 1 000 раз) под влиянием диуретического гормона, секретируемого в грудных ганглиях. Однако его выведение в гемолимфу происходит только при возбуждении рецепторов растяжения брюшка, что наблюдается всякий раз при насасывании крови. У саранчи Schistocerca gregaria Forsk. диуретический гормон стимулирует абсорбцию в мальпигиевых сосудах и тормозит реабсорбцию в ректальных железах задней кишки. У таракана Periplaneta americana L. наряду с диуретическим выделяется антидиуретический гормон.
Кроме мальпигиевых сосудов функции экскреции конечных продуктов метаболизма азота выполняют лабиальные железы Collembola, Thysanura и некоторых крылатых насекомых. У шелкопряда Hyalophora cecropia L. лабиальные шёлкоотделительные железы гусениц преобразуются в имагинальные органы, регулирующие водообмен и выделение экскретов. Продуцируемая придаточными половыми железами самцов некоторых тараканов мочевая кислота используется для покрытия сперматофоров и таким образом выводится из организма. Вместе с тем азотсодержащие метаболиты часто вообще не выводятся наружу, а, накапливаясь в уратных клетках жирового тела, в нефроцитах и в кутикуле, исключаются из процессов обмена веществ.
Согласованность и совершенство рассмотренных процессов метаболизма обеспечивают экономное расходование воды и энергетических субстратов, не допуская потерь сколько-нибудь ценных метаболитов. В этом отношении насекомые не уступают млекопитающим животным, несмотря на то что малые размеры тела определяют для них ряд ограничений. Однако ключевые пути метаболизма у тех и других принципиально сходны.
Источник