
Какие продукты и сколько молекул атф получаются при гликолизе

Синтез АТФ при расщеплении глюкозы. Выделение энергии из гликогена
Мы можем определить общее количество молекул АТФ, которое образуется при расщеплении 1 молекулы глюкозы при оптимальных условиях.
1. Во время гликолиза образуются 4 молекулы АТФ: 2 молекулы АТФ расходуются на первом этапе фосфорилирования глюкозы, необходимого для хода процесса гликолиза, чистый выход АТФ при гликолизе равен 2 молекулам АТФ.
2. В итоге цикла лимонной кислоты образуется 1 молекула АТФ. Однако в связи с тем, что 1 молекула глюкозы расщепляется на 2 молекулы пировиноградной кислоты, каждая из которых проходит оборот в цикле Кребса, получается чистый выход АТФ на 1 молекулу глюкозы, равный 2 молекулам АТФ.
3. При полном окислении глюкозы суммарно образуются 24 атома водорода в связи с процессом гликолиза и циклом лимонной кислоты, 20 из них окисляются в соответствии с хемо-осмотическим механизмом с выделением 3 молекул АТФ на каждые 2 атома водорода. В итоге получается еще 30 молекул АТФ.
4. Четыре оставшихся атома водорода выделяются под влиянием дегидрогеназ и включаются в цикл хемоосмотического окисления в митохондриях помимо первой стадии. Окисление 2 атомов водорода сопровождается получением 2 молекул АТФ, в итоге получается еще 4 молекулы АТФ.
Сложив все полученные молекулы, получим 38 молекул АТФ как максимально возможное количество при окислении 1 молекулы глюкозы до углекислого газа и воды. Следовательно, 456000 калорий могут сохраняться в виде АТФ из 686000 калорий, получаемых при полном окислении 1 грамм-молекулы глюкозы. Эффективность преобразования энергии, обеспечиваемая этим механизмом, составляет около 66%. Остальные 34% энергии преобразуются в тепловую и не могут быть использованы клетками для выполнения специфических функций.
Выделение энергии из гликогена
Продолжительное высвобождение энергии из глюкозы, когда клетки не нуждаются в энергии, было бы слишком расточительным процессом. Гликолиз и последующее окисление атомов водорода постоянно контролируются в соответствии с потребностями клеток в АТФ. Этот контроль осуществляется многочисленными вариантами управляющих механизмов обратной связи в ходе химических реакций. К числу наиболее важных влияний такого рода можно отнести концентрацию АДФ и АТФ, контролирующую скорость химических реакций в ходе процессов обмена энергии.
Одним из важных путей, позволяющих АТФ управлять обменом энергии, является ингибирование фермента фосфофруктокиназы. Этот фермент обеспечивает образование фруктозо-1,6-дифосфата — одной из начальных стадий гликолиза, поэтому результирующим влиянием избытка АТФ в клетке будет торможение или даже остановка гликолиза, что, в свою очередь, приведет к торможению обмена углеводов. АДФ (равно как и АМФ) оказывает противоположное влияние на фосфофруктокиназу, существенно повышая ее активность. Когда АТФ используется тканями для энергообеспечения большинства химических реакций в клетках, это уменьшает ингибирование фермента фосфофруктокиназы, более того, его активность повышается параллельно увеличению концентрации АДФ. В результате запускаются процессы гликолиза, приводящие к восстановлению запасов АТФ в клетках.
Другой способ управления опосредован цитратами, образующимися в цикле лимонной кислоты. Избыток этих ионов существенно снижает активность фосфофруктокиназы, что не дает гликолизу опережать скорость использования пировиноградной кислоты, образующейся в результате гликолиза в цикле лимонной кислоты.
Третий способ, с помощью которого система АТФ-АДФ-АМФ может контролировать обмен углеводов и управлять выделением энергии из жиров и белков, заключается в следующем. Возвращаясь к различным химическим реакциям, служащим способом выделения энергии, мы можем заметить, что если весь имеющийся в наличии АМФ уже превращен в АТФ, дальнейшее образование АТФ становится невозможным. В результате прекращаются все процессы использования питательных веществ (глюкозы, белков и жиров) для получения энергии в виде АТФ. Лишь после использования образовавшегося АТФ в качестве источника энергии в клетках для обеспечения разнообразных физиологических функций вновь появляющиеся АДФ и АМФ запустят процессы получения энергии, в ходе которых АДФ и АМФ преобразуются в АТФ. Этот путь автоматически поддерживает сохранение определенных запасов АТФ, кроме случаев экстремальной активности клеток, например при тяжелых физических нагрузках.
– Также рекомендуем “Анаэробный гликолиз. Молочная и пировиноградная кислота”
Оглавление темы “Синтез АТФ. Обмен глюкозы и жиров”:
1. Образование АТФ через хемоосмотический механизм. Образование и синтез АТФ
2. Синтез АТФ при расщеплении глюкозы. Выделение энергии из гликогена
3. Анаэробный гликолиз. Молочная и пировиноградная кислота
4. Высвобождение энергии из глюкозы через пентозофосфатный цикл. Превращение глюкозы в жиры
5. Образование углеводов из белков и жиров. Регуляция глюконеогенеза
6. Метаболизм жиров в организме. Транспорт липидов
7. Свободные жирные кислоты. Транспорт свободных жирных кислот
8. Липопротеины. Виды и физиология липопротеинов
9. Обмен жиров и их депонирование. Жиры печени
10. Расщепление жирных кислот. Окисление жирных кислот
Источник
Гликолиз – процесс окисления глюкозы, при котором из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты, не является мембранозависимым процессом. Он происходит в цитоплазме. Однако ферменты гликолиза связаны со структурами цитоскелета. Суть гликолиза состоит в том, что молекула глюкозы (C6H12O6) без участия кислорода распадается на две молекулы пировиноградной кислоты (СН3СОСООН). При этом окисление идет за счет отщепления от молекулы глюкозы четырех атомов водорода, связывающихся со сложным органическим веществом НАД с получением двух молекул НАД•Н. Выделяющаяся при этом энергия запасается (40% от общего количества) в виде макроэргических связей двух молекул АТФ. 60% энергии выделяется в виде тепла. При последующем окислении НАД•Н получается еще 6 молекул АТФ. Таким образом, полный энергетический выход гликолиза в анаэробных условиях составляет 8 молекул АТФ.
На схеме в рамках обозначены исходные субстраты и конечные продукты гликолиза, цифрами в скобках – число молекул.
Для распада и частичного окисления молекулы глюкозы требуется протекание 11 сложных последовательных реакций.
Реакции гликолиза
Ход реакций
Ферменты, Активаторы, ингибиторы
Подготовительная стадия гликолиза
Стадия активации глюкозы проходит в 5 реакций, в ходе которых 1 молекула гексозы (глюкозы) расщепляется на 2 молекулы триоз-глицеральдегидфосфата
1. Необратимая реакция фосфорилирования глюкозы
Процесс гликолиза начинается с фосфорилирования глюкозы за счет АТФ – первая реакция. Это первая пусковая реакция гликолиза. Ее результатом является глюкозо-6-фосфат, имеющий отрицательный заряд. В гликолизе может участвовать не только глюкоза, но и другие гексозы (фруктоза), но в результате фосфорилирования и активации все равно образуется глюкозо-6-фосфат.

фермент: гексокиназа
Активаторы: АДФ, Н3РO4.
Ингибиторы: глюкозо-6-Ф, фосфоенолпируват.
2. Обратимая реакция изомеризации глюкозо-6-фосфата
Во второй реакции происходит изомеризация (внутримолекулярные перестройки) глюкозо-6-фосфата во фруктозо-6-фосфат.

фермент: глюкозо-6-фосфатизомераза
3. Необратимая реакция фосфорилирования фруктозо-6-фосфата (ключевая стадия гликолиза)
В третьей реакции происходит фосфорилирование (присоединение остатка ортофосфорной кислоты) фруктозо-6-фосфата с образованием фруктозо-1,6-дифосфата. При этом затрачивается еще одна молекула АТФ (уже вторая) – это вторая пусковая реакция гликолиза. Она идет в присутствии Mg2+ и является необратимой, так как сопровождается масштабным уменьшением свободной энергии.

фермент: фосфофруктокиназа
Активаторы: АДФ, АМФ, Н3РO4, К+.
Ингибиторы: АТФ, цитрат, НАДН.
4. Обратимая реакция дихотомического расщепления фруктозо-1,6-дифосфата
В четвертой реакции гликолиза происходит расщепление фруктозо-1,6-дифосфата на две молекулы глицеральдегид-3-фосфата.
фермент: алъдолаза
5. Обратимая реакция изомеризации дигидроксиацетона-3-фосфат в глицеральдегид-3-фосфат
В пятой реакции происходит изомеризация полученных триозофосфатов. На этом заканчивается первая стадия гликолиза.

фермент: триозофосфатизомераза
Стадия генерации АТФ
Проходит в 6 реакций (или 5), в ходе которых энергия окислительных реакций трансформируется в химическую энергию АТФ (субстратное фосфорилирование).
6. Окисление глицеральдегид-3-фосфата до 1,3-дифосфоглицерата (реакция гликолитической оксиредукции)
В шестой реакции происходит окисление альдегидной группы до карбоксильной. Выделившийся Н+ акцептируется NAD, который восстанавливается до NADH. Освобождающаяся энергия затрачивается для образования высокоэнергетической связи 1,3-бифосфоглицерата (1,3-бифосфоглицериновая кислота).

фермент: глицералъдегид-3-фосфат-дегидрогеназа
7. Субстратное фосфорилирование АДФ (7)
В седьмой реакции фосфорильная группа 1,3-бифосфоглицерата переносится на ADP, в результате чего образуется АТР (напоминаем, что следует иметь в виду две параллельные цепи реакций, с участием двух молекул триоз, образовавшихся из одной молекулы гексозы, следовательно, синтезируется не одна, а две молекулы АТР).

фермент: фосфоглицераткиназа
8. Реакция изомеризации 3-фосфоглицерата в 2-фосфоглицерат
В восьмой реакции гликолиза происходит перенос фосфатной группы с третьего атома углерода на второй. В результате образуется 2-фосфоглицерат (2-фосфоглицериновая кислота).

9. Реакция енолизации
Девятая реакция сопровождается внутримолекулярными окислительно-восстановительными процессами, в результате которых образуется фосфоенолпируват (фосфоенолпировиноградная кислота) с высокоэнергетической связью во втором атоме углерода и отщепляется молекула воды

фермент: енолаза
10. Реакция субстратного фосфорилирования
В ходе десятой реакции фосфорильная группа переносится на ADP. При этом синтезируется АТР и пируват (пировиноградная кислота). Эта реакция также необратима, поскольку высокоэкзергонична.

фермент: пируваткиназа
11. Реакция обратимого восстановления пировиноградной кислоты до молочной кислоты (в анаэробных условиях)
Если после гликолиза следует аэробное расщепление, пируват мигрирует в матрикс митохондрий, где, взаимодействуя с коэнзимом-А, участвует в образовании ацетил-СоА. В анаэробных условиях пируват при участии NADH восстанавливается до лактата (молочной кислоты), который при этом является конечным продуктом гликолиза. Затем в аэробных условиях лактат может обратно превратиться в пируват и окислиться в митохондриях.

фермент: лактатдегидрогеназа
1. Биология для поступающих в вузы / Г.Л. Билич, В.А. Крыжановский. — 2008.
2. Биология в таблицах и схемах / Спб. — 2004.
3. Биохимия в схемах и таблицах / И. В. Семак – Минск — 2011.
Источник
Энергетический обмен
Энергетический обмен (катаболизм, диссимиляция) — совокупность реакций расщепления органических веществ, сопровождающихся выделением энергии. Энергия, освобождающаяся при распаде органических веществ, не сразу используется клеткой, а запасается в форме АТФ и других высокоэнергетических соединений. АТФ — универсальный источник энергообеспечения клетки. Синтез АТФ происходит в клетках всех организмов в процессе фосфорилирования — присоединения неорганического фосфата к АДФ.
У аэробных организмов (живущих в кислородной среде) выделяют три этапа энергетического обмена: подготовительный, бескислородное окисление и кислородное окисление; у анаэробных организмов (живущих в бескислородной среде) и аэробных при недостатке кислорода — два этапа: подготовительный, бескислородное окисление.
Подготовительный этап
Заключается в ферментативном расщеплении сложных органических веществ до простых: белковые молекулы — до аминокислот, жиры — до глицерина и карбоновых кислот, углеводы — до глюкозы, нуклеиновые кислоты — до нуклеотидов. Распад высокомолекулярных органических соединений осуществляется или ферментами желудочно-кишечного тракта или ферментами лизосом. Вся высвобождающаяся при этом энергия рассеивается в виде тепла. Образовавшиеся небольшие органические молекулы могут быть использованы в качестве «строительного материала» или могут подвергаться дальнейшему расщеплению.
Бескислородное окисление, или гликолиз
Этот этап заключается в дальнейшем расщеплении органических веществ, образовавшихся во время подготовительного этапа, происходит в цитоплазме клетки и в присутствии кислорода не нуждается. Главным источником энергии в клетке является глюкоза. Процесс бескислородного неполного расщепления глюкозы — гликолиз.
Потеря электронов называется окислением, приобретение — восстановлением, при этом донор электронов окисляется, акцептор восстанавливается.
Следует отметить, что биологическое окисление в клетках может происходить как с участием кислорода:
А + О2 → АО2,
так и без его участия, за счет переноса атомов водорода от одного вещества к другому. Например, вещество «А» окисляется за счет вещества «В»:
АН2 + В → А + ВН2
или за счет переноса электронов, например, двухвалентное железо окисляется до трехвалентного:
Fe2+ → Fe3+ + e—.
Гликолиз — сложный многоступенчатый процесс, включающий в себя десять реакций. Во время этого процесса происходит дегидрирование глюкозы, акцептором водорода служит кофермент НАД+ (никотинамидадениндинуклеотид). Глюкоза в результате цепочки ферментативных реакций превращается в две молекулы пировиноградной кислоты (ПВК), при этом суммарно образуются 2 молекулы АТФ и восстановленная форма переносчика водорода НАД·Н2:
С6Н12О6 + 2АДФ + 2Н3РО4 + 2НАД+ → 2С3Н4О3 + 2АТФ + 2Н2О + 2НАД·Н2.
Дальнейшая судьба ПВК зависит от присутствия кислорода в клетке. Если кислорода нет, у дрожжей и растений происходит спиртовое брожение, при котором сначала происходит образование уксусного альдегида, а затем этилового спирта:
- С3Н4О3 → СО2 + СН3СОН,
- СН3СОН + НАД·Н2 → С2Н5ОН + НАД+.
У животных и некоторых бактерий при недостатке кислорода происходит молочнокислое брожение с образованием молочной кислоты:
С3Н4О3 + НАД·Н2 → С3Н6О3 + НАД+.
В результате гликолиза одной молекулы глюкозы высвобождается 200 кДж, из которых 120 кДж рассеивается в виде тепла, а 80% запасается в связях АТФ.

Кислородное окисление, или дыхание
Заключается в полном расщеплении пировиноградной кислоты, происходит в митохондриях и при обязательном присутствии кислорода.
Пировиноградная кислота транспортируется в митохондрии (строение и функции митохондрий — лекция №7). Здесь происходит дегидрирование (отщепление водорода) и декарбоксилирование (отщепление углекислого газа) ПВК с образованием двухуглеродной ацетильной группы, которая вступает в цикл реакций, получивших название реакций цикла Кребса. Идет дальнейшее окисление, связанное с дегидрированием и декарбоксилированием. В результате на каждую разрушенную молекулу ПВК из митохондрии удаляется три молекулы СО2; образуется пять пар атомов водорода, связанных с переносчиками (4НАД·Н2, ФАД·Н2), а также одна молекула АТФ.

Суммарная реакция гликолиза и разрушения ПВК в митохондриях до водорода и углекислого газа выглядит следующим образом:
С6Н12О6 + 6Н2О → 6СО2 + 4АТФ + 12Н2.
Две молекулы АТФ образуются в результате гликолиза, две — в цикле Кребса; две пары атомов водорода (2НАДЧН2) образовались в результате гликолиза, десять пар — в цикле Кребса.
Последним этапом является окисление пар атомов водорода с участием кислорода до воды с одновременным фосфорилированием АДФ до АТФ. Водород передается трем большим ферментным комплексам (флавопротеины, коферменты Q, цитохромы) дыхательной цепи, расположенным во внутренней мембране митохондрий. У водорода отбираются электроны, которые в матриксе митохондрий в конечном итоге соединяются с кислородом:
О2 + e— → О2—.
Купить проверочные работы
и тесты по биологии




Протоны закачиваются в межмембранное пространство митохондрий, в «протонный резервуар». Внутренняя мембрана непроницаема для ионов водорода, с одной стороны она заряжается отрицательно (за счет О2—), с другой — положительно (за счет Н+). Когда разность потенциалов на внутренней мембране достигает 200 мВ, протоны проходят через канал фермента АТФ-синтетазы, образуется АТФ, а цитохромоксидаза катализирует восстановление кислорода до воды. Так в результате окисления двенадцати пар атомов водорода образуется 34 молекулы АТФ.

1 — наружная мембрана; 2 — межмембранное пространство, протонный резервуар;
3 — цитохромы; 4 — АТФ-синтетаза.
При перфорации внутренних митохондриальных мембран окисление НАД·Н2 продолжается, но АТФ-синтетаза не работает и образования АТФ в дыхательной цепи не происходит, энергия рассеивается в форме тепла (клетки «бурого жира» млекопитающих).
Суммарная реакция расщепления глюкозы до углекислого газа и воды выглядит следующим образом:
С6Н12О6 + 6О2 → 6СО2 + 6Н2О + 38АТФ + Qт,
где Qт — тепловая энергия.
Перейти к лекции №10 «Понятие об обмене веществ. Биосинтез белков»
Перейти к лекции №12 «Фотосинтез. Хемосинтез»
Смотреть оглавление (лекции №1-25)
Источник
ГЛИКОЛИЗ. ГЛЮКОНЕОГЕНЕЗ. АЭРОБНОЕ ОКИСЛЕНИЕ ГЛЮКОЗЫ.
ГЛИКОЛИЗ
Гликолиз — это сложный ферментативный процесс расщепления глюкозы до двух молекул пирувата (аэробный гликолиз) или двух молекул лактата (анаэробный гликолиз, протекающий без потребления кислорода).
Суммарное уравнение анаэробного гликолиза:
![]()
Гликолиз функционирует во всех живых клетках. Все ферменты локализованы в цитозоле, формируя полиферментный комплекс.
Гликолиз осуществляется в два этапа.
I. Подготовительный этап — дихотомический распад глюкозы на две молекулы глицеральдегид-3-фосфата. Превращения сопровождаются затратой 2 АТФ.
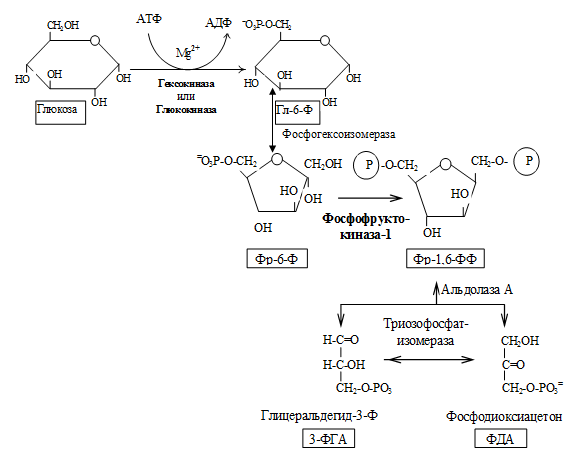
II. Этап гликолитической оксидоредукции — превращение глицеральдегид-3-фосфата в лактат. Включает окислительно-восстановительные реакции и реакции фосфорилирования, сопровождающиеся генерацией АТФ.
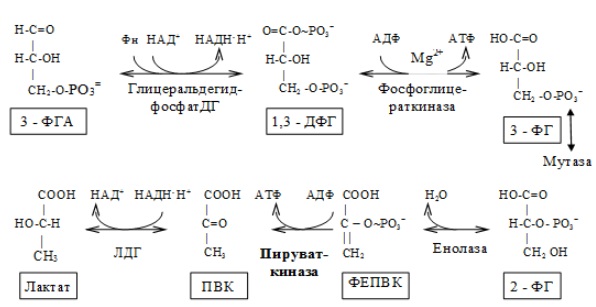
На втором этапе окисляются две молекулы глицеральдегид-3-фосфата, поэтому в реакциях впереди формулы субстрата следует ставить коэффициент 2.
В анаэробных условиях окисление НАДН.Н+, восстановленного в глицеральдегидфосфатдегидрогеназной реакции, происходит в лактатдегидрогеназной реакции. В аэробных условиях НАДН.Н+ окисляется кислородом с участием ферментов дыхательной цепи, а выделяющаяся при этом энергия используется на синтез 1,5 или 2,5 моль АТФ (в зависимости от челночного механизма транспорта гликолитического НАДН.Н+ в митохондрию).
Энергетический баланс гликолиза — две молекулы АТФ на одну молекулу глюкозы. На I этапе гликолиза расходуются две молекулы АТФ для активирования субстрата (в гексокиназной и фосфофруктокиназной реакциях). На II этапе образуются четыре молекулы АТФ (в фосфоглицераткиназной и пируваткиназной реакциях). Синтез АТФ осуществляется путем субстратного фосфорилирования.
Ключевые ферменты гликолиза:
1. Гексокиназа — это регуляторный фермент гликолиза во внепеченочных клетках. Гексокиназа аллостерически ингибируется глюкозо-6-фосфатом. Глюкокиназа — регуляторный фермент гликолиза в гепатоцитах. Синтез глюкокиназы индуцируется инсулином.
2. Фосфофруктокиназа-1. Это главный ключевой фермент, катализирует реакцию, лимитирующую скорость всего процесса (наиболее медленная реакция). Синтез фермента индуцируется инсулином. Аллостерические активаторы — АМФ, АДФ, фруктозо-2,6-дифосфат. Уровень фруктозо-2,6-дифосфата увеличивается под действием инсулина и понижается под действием глюкагона. Аллостерические ингибиторы — АТФ, цитрат.
3. Пируваткиназа. Фермент активен в нефосфорилированной форме. Глюкагон (в гепатоцитах) и адреналин (в миоцитах) стимулируют фосфорилирование фермента, а значит инактивируют фермент. Инсулин, наоборот, стимулирует дефосфорилирование фермента, а значит активирует фермент. Аллостерический активатор — Фр-1,6-ФФ. Аллостерический ингибитор — АТФ, ацетилКоА. Синтез фермента индуцирует инсулин.
Биологическая роль гликолиза:
1. Генерирование АТФ. Гликолиз — единственный процесс в клетках, продуцирующий АТФ без потребления кислорода. Клетки, имеющие мало или не имеющие вообще митохондрий, получают АТФ только в ходе гликолиза.
Значение гликолиза для эритроцитов. Гликолиз — единственный процесс, продуцирующий АТФ в эритроцитах и поддерживающий их целостность и функции.
Наследственный дефект пируваткиназы сопровождается гемолитической анемией. При этой патологии эритроциты имеют от 5 до 25 % нормальной пируваткиназной активности и, следовательно, скорость гликолиза низкая.
Промежуточный продукт гликолиза в эритроцитах — 2,3-дифосфоглицерат (2,3-ДФГ) — понижает сродство гемоглобина к кислороду, способствуя диссоциации кислорода из оксигемоглобина и переходу его в ткани. Нарушения гликолиза в эритроцитах могут оказывать влияние на транспорт кислорода. Так, при недостаточности гексокиназы наблюдается понижение уровня 2,3-ДФГ и ненормально высокое сродство гемоглобина к кислороду. И наоборот, при недостаточности пируваткиназы содержание 2,3-ДФГ вдвое превышает норму, что обусловливает низкое сродство гемоглобина к кислороду.
2. Является источником углеводородных радикалов для процессов биосинтеза в клетках:
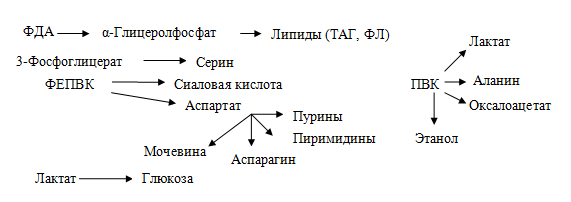
Патогенетическая взаимосвязь углеводов пищи и кариеса
Потребление легкоферментируемых углеводов, в частности сахарозы, инициирует кислотную деминерализацию эмали зубов. У бактерий имеются два альтернативных пути использования пирувата: первый — путь восстановления ПВК в молочную кислоту с участием лактатдегидрогеназы (ЛДГ), второй — расщепление ПВК на уксусную и муравьиную кислоту с участием пируватформиатлиазы (ПФЛ).
Схема расщепления сахарозы под влиянием ферментов бактерий полости рта:
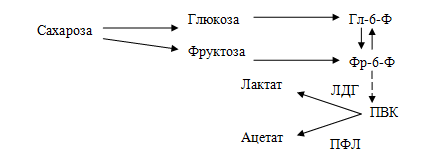
Зависимость выхода Са2+ из зубов от рН ротовой жидкости:
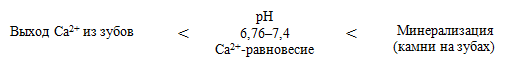
Аэробное окисление глюкозы
Это основной путь катаболизма глюкозы у аэробных организмов. Процесс осуществляется в три этапа. В аэробных условиях глюкоза окисляется до СО2 и Н2О.
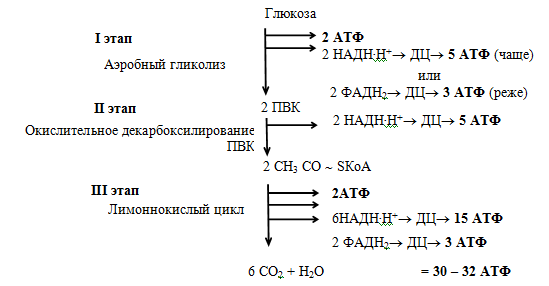
Энергетический баланс. Энергетический баланс аэробного окисления глюкозы —
30–32 моля АТФ на молекулу глюкозы.
ГЛЮКОНЕОГЕНЕЗ
Глюконеогенез — синтез глюкозы из соединений неуглеводной природы.
В организме взрослого человека за сутки может синтезироваться до 250 г глюкозы.
Глюконеогенез осуществляется главным образом в печени (синтезируетя до 90 % всей глюкозы), в корковом веществе почек и в энтероцитах (совсем незначительно).
Глюконеогенез стимулируется при длительном голодании, при ограничении поступления углеводов с пищей, в период восстановления после мышечной нагрузки, у новорождённых в первые часы после рождения.
Субстраты глюконеогенеза. Истинными субстратами глюконеогенеза являютя пируват, оксалоацетат, фосфодиоксиацетон, которые непосредственно включаются в этот процесс. Все вещества неуглеводной природы, дающие эти метаболиты, являются субстратами глюконеогенеза: лактат→ПВК, метаболиты цикла Кребса→ЩУК, глицерол→фосфодиоксиацетон, пропионил-КоА→метаболиты цикла Кребса→ЩУК, глюкогенные аминокислоты→ПВК или ЩУК. Главный источник субстратов глюконеогенеза — глюкогенные аминокислоты. К глюкогенным аминокислотам относятся все протеиногенные аминокислоты, кроме лейцина и лизина.
Стехиометрия:
2ПВК + 4АТФ + 2ГТФ + 2НАДН.Н+ + 2Н + 6Н2О → Глюкоза + 4АДФ + 2ГДФ + 6Фн + 2НАД +
Глюконеогенез протекает, в основном, по тому же пути, что и гликолиз, но в обратном направлении. Для обхода трех ключевых реакций гликолиза используются четыре специфических фермента глюконеогенеза.
Ключевые ферменты и ключевые реакции глюконеогенеза:
1. Пируваткарбоксилаза

2. Фосфоенолпируваткарбоксикиназа
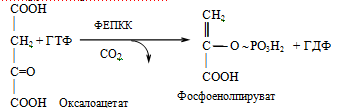
3. Фруктозо-1,6-бисфосфатаза
Фруктозо-1,6-бисфосфат + Н2О Фруктозо-6-фосфат + ФН
4. Глюкозо-6-фосфатаза
Глюкозо-6-фосфат + Н2О Глюкоза + ФН
Энергетический баланс. На синтез молекулы глюкозы из двух молекул пирувата расходуется 4АТФ и 2ГТФ (6АТФ). Энергию для глюконеогенеза поставляет процесс β-окис-ления жирных кислот.
Регуляция глюконеогенеза.
Глюконеогенез стимулируется в условиях гипогликемии при низком уровне инсулина и преобладании его антагонистов (глюкагона, катехоламинов, глюкокортикоидов).
1. Регуляция активности ключевых ферментов:
- фруктозо-1,6-бисфосфатаза по аллостерическому механизму активируется АТФ, ингибирутся Фр-1,6-ФФ и АМФ;
- пируваткарбоксилаза активируется СН3СО~КоА (аллостерический активатор).
2. Регуляция количества ключевых ферментов: глюкокортикоиды и глюкагон индуцируют синтез ключевых ферментов, а инсулин — репрессирует.
3. Регуляция количества субстрата: количество субстратов глюконеогенеза увеличивается под действием глюкокортикоидов (катаболическое действие на белки мышечной и лимфоидной ткани, на жировую ткань), а также глюкагона (катаболическое действие на жировую ткань).
Биологическая роль глюконеогенеза:
1. Поддержание уровня глюкозы в крови. При длительном голодании (голодание более суток) глюконеогенез является единственным процессом, поставляющим глюкозу в кровь.
2. Возвращение лактата в метаболический фонд углеводов. Лактат, образующийся в процессе анаэробного окисления глюкозы в эритроцитах и скелетных мышцах, транспортируется кровью в печень и превращается в гепатоцитах в глюкозу. Это так называемый межорганный цикл Кори.
3. Предотвращение лактатного ацидоза, то есть в ходе глюконеогенеза лактат крови превращается в глюкозу.
Источник